В якому варіанті правильно і найповніше перелічені елементи жіночої статевої системи ?
Оберіть найповніше визначення — «Фагоцитоз — це …»
a)процес, важливий у реакції організму на інфекцію
b)процес, властивий найпростішим, кишковопорожни
…
нним, а також клітинам крові — лейкоцитам
c)процес активного поглинання і внутрішньоклітинного перетравлення живих і неживих частинок одноклітинними організмами або окремими клітинами багатоклітинних тваринних організмів
please help me Біологія
5. Як називається процес виникнення нових структур і зміна їх форми в процесі
онтогенезу:
A. морфогенез
B. органогенез
C. гістогенез
D. гаструляція
E.
…
запліднення
12.Період дроблення зиготи відрізняється від інших видів поділу клітин:
A.інтенсивним ростом дочірніх клітин;
B.відсутністю росту дочірніх клітин;
C.редукцією хромосом;
D.кон’югацією хромосом;
E.додатковими стадіями мітозу
13.Зародковий розвиток – це початковий період:
A.філогенезу;
B.дроблення;
C.
овогенезу;
D.кон’югації хромосом;
E.онтогенезу
14.Нервова трубка і хорда в зародка формується з:
A.ектодерми;
B.ентодерми;
C.мезодерми;
D.нейрули;
E.бластули
16.У рослин усі клітини утворюються з:
A.ектодерми;
B.ентодерми;
C.мезодерми;
D.нейрули;
E. твірної меристеми
17.Ембріогенез закінчується на стадії гаструли в:
A.ланцетника;
B.гідри;
C.дощового черв’яка;
D. інфузорії туфельки;
E. малярійного плазмодія
18.Морула – це стадія:
A.ембріогенезу;
B.органогенезу;
C.гістогенезу;
D.цитокінезу;
E. овогенезу
22.Яйцеклітина в покритонасінних рослин розвивається в:
A.пиляках;
B.зав’язі;
C.архегоніях;
D.спорах;
E. тичинках
25.Назвіть етапи онтогенезу:
A.пренатальний,овогенез;
B. пренатальний,сперматогенез;
C.постнатальний,овогенез;
D.пренатальний,постнатальний;
E.постнатальний,сперматогенез;
26.Назвіть періоди пренатального онтогенезу:
A.початковий,зародковий;
B.початковий,плодовий;
C.немає періодів;
D. початковий,зародковий,плодовий;
E.початковий,проміжний,кінцевий;
31.Вкажіть,яку функцію не виконує плацента?:
A. трофічну;
B.захисну;
C.ендокринну;
D.імунну;
E. кровотворну
32.Що таке нейрула?:
A.нервова трубка;
B.зародок на стадії утворення нервової трубки;
C.нервові гребені;
D.гангліозні пластинки;
E. порожнина нервової трубки
33.З яких клітин складається бластула
A. Трофобластів.
B. Фолікулярних.
C. Децидуальних.
D.Бластомерів
E. Амніотичних.
35.З якої складової частини бластоцисти утворюється хоріон?
A. Трофобласта
B. Бластоцелі.
C. Амніобласта.
D. Ембріобласта.
E. Бластомерів.
37.Основна біологічна роль яйцеклітини в процесі запліднення:
A. Зустріч із сперматозоїдом.
B. Доповнення диплоїдного набору хромосом, трофічна.
C. Визначення статі зародка
D. Забезпечення моноспермії.
E. Захисна.
38.Основні структурні компоненти головки сперматозоїда:
A. Диплоїдне ядро і акросома.
B. Гаплоїдне ядро і мітохондрії.
C. Диплоїдне ядро і мітохондрії.
D. Гаплоїдне ядро і акросома.
E. Гаплоїдне ядро, акросома і мітохондрії.
39. Перебуваючи на стадії дроблення зародок побудований з клітин:
A. Бластомерів.
B. Трофобластів.
C. Цитотрофобластів.
D. Ембріобластів.
E. Бластоцелей.
40.Сперматозоїди людини містять наступні відділи:
A. Головку, аксонему, хвостик.
B. Головку, хвостик, ядро, шийку.
C. Акросому, ядро, хвостик.
D. Головку, шийку, хвостик.
E. Акросому, аксонему, хвостик.
1. Виберіть приклад, який ілюструє внутрiшньовидову гібридизацію: А) схрещування яблуні та груші Б) схрещування білуги та стерлядi В) отримання курей-
…
бройлерів Г) отримання поліплоїдних сортів 2. Що таке біотехнологія? А) галузь генетики, яка виводить в та поліпшує існуючі сорти рослин і породи i тварин Б) галузь цитологiї В) галузь біохімії Г) сукупність промислових методів, у яких використовують органiзми чи біологічні процеси Назвiть чинник ,що є рушійною силою селекцiйного процесу 3. А) мутації Б) рекомбінації В) штучний добір Г) гібридизація 4. Вкажіть центр походження картоплі А) Південно-азійський Б) Середземноморський в Абiссiнський Г)АНДIЙСЬКИЙ
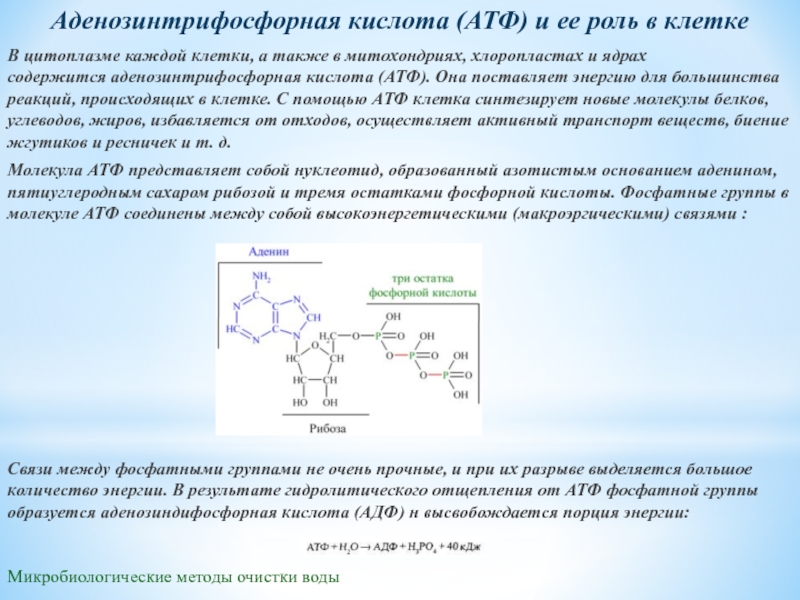
§ 8. Атф. Биологически активные вещества
АТФ. Нуклеотиды являются структурной основой
для синтеза целого ряда важных для
жизнедеятельности органических веществ.
Наиболее широко распространенными
среди них являются макроэргические
соединения (вещества, содержащие богатые
энергией, или макроэргические связи).
К таким веществам в первую очередь
относится аденозинтрифосфорная
кислота (АТФ). АТФ – универсальный
хранитель и переносчик энергии в клетках
всех организмов. Практически все идущие
с затратой энергии биохимические
процессы в клетках организмов в качестве
ее источника используют АТФ. С
использованием энергии АТФ в клетке
синтезируют новые молекулы белков,
углеводов, жиров, осуществляется
активный транспорт веществ, биение
жгутиков и ресничек, поддерживается
постоянная температура тела теплокровных
организмов и т.д.
Молекула
АТФсостоит
из азотистого основания аденина,
пятиуглеродного сахара рибозы и трех остатков фосфорной
кислоты.
Фосфатные группы в молекуле АТФ соединены
между собой высокоэнергетическими
(макроэргическими) связями (в формуле
обозначены символом ~) (рис. ).
Связи
между фосфатными группами не очень
прочные, и при их разрыве выделяется
большое количество энергии. В
результате гидролитического отщепления
от АТФ фосфатной группы образуется аденозиндифосфорная
кислота (АДФ) и высвобождается порция энергии:
АТФ
+ Н2О
→ АДФ + Н3РО 4 + 40 кДж.
АДФ может подвергаться
дальнейшему гидролизу с отщеплением
еще одной фосфатной группы и выделением
второй порции энергии. При этом АДФ
преобразуется в аденозинмонофосфат
(АМФ).
АДФ
+ Н2 О
→ АМФ + Н3Р04 + 40 кДж.
Обратный
процесс — синтез АТФ происходит в
результате присоединения к молекуле
АДФ остатка фосфорной кислоты
(реакция фосфорилирования). Этот
процесс осуществляется за счет
энергии, освобождающейся при окислении
органических веществ (окислительное
фосфорилирование). Для
образования 1 моля АТФ должно быть
затрачено не менее 40 кДж энергии, которая
аккумулируется в ее макроэргических
связях:
АДФ
+ Н3Р04 + 40 кДж → АТФ + Н20.
АТФ чрезвычайно
быстро обновляется. У человека,
например, каждая молекула АТФ расщепляется
и вновь восстанавливается 2400 раз в
сутки, так что средняя продолжительность
ее жизни менее 1 мин. Синтез АТФ
осуществляется, главным образом, в
митохондриях и хлоропластах, частично
в цитоплазме.
Макроэргические
соединения могут образовываться и на
основе других нуклеотидов. Например,
гуанозинтрифосфат (ГТФ) играет важную
роль в ряде биохимических процессов.
Однако АТФ является наиболее
распространенным и универсальным
источником энергии в клетках живых
организмов.
Биологически
активные вещества. Особой
группой органических соединений живых
организмов являются биологически
активные вещества. Они регулируют
процессы обмена веществ, роста и развития
организмов, служат для защиты или влияют
на особей своего или других видов.
Одной
из групп биологически активных
веществ являются витамины. Витамины —
это низкомолекулярные органические
вещества разнообразного строения , необходимые
для жизнедеятельности всех живых
организмов. Они принимают участие в
обмене веществ и превращении энергии,
преимущественно как компоненты сложных
ферментов.
Традиционно
витамины обозначают буквами латинского
алфавита А, В, С, D
и т.д., но у каждого из них есть и названия.
Например, витамин С — аскорбиновая
кислота, витамин А – ретинол и так далее.
Одни
витамины растворяются в жирах и их
называют жирорастворимыми (A,
D,
E,
K),
другие растворимы в воде и соответственно
называются водорастворимыми (С,
В, РР, Н).
Сейчас
известно около 50 различных витаминов
и витаминоподобных веществ. Они по-разному
влияют на живые организмы, однако
являются жизненно необходимыми
компонентами сбалансированного питания
человека и животных. Основным источником
витаминов для человека и животных
являются продукты питания преимущественно
растительного происхождения. Однако
некоторые витамины содержатся только
в продуктах животного происхождения
(например, витамины А и D).
Некоторые витамины могут в небольшом
количестве синтезироваться в организме
человека и животных из предшественников
(провитаминов). Например, в коже человека
под действием ультрафиолетового
солнечного излучения синтезируется
витамин D.
Витамины в организме человека и животных
могут синтезировать симбиотические
микроорганизмы. Например, в кишечнике
человека они синтезируют витамины К,
В
,
В
.
Однако образованных в организме человека
витаминов недостаточно для обеспечения
его нормальной жизнедеятельности.
Необходимо их дополниельное поступление
извне, с пищей.
При
недостатке в организме витаминов,
развивается гиповитаминоз(от
греч. гипо
— под,
ниже), при полном их отсутствии — авитаминоз, а
при избытке — гипервитаминоз(от
греч. гипер
— сверх).
Гипо- и авитаминоз могут возникнуть
и вследствие нарушения обмена веществ,
когда организм не воспринимает некоторые
витамины.
Гормоны (отгреч.гормао
— двигаю,
побуждаю) — органические вещества, способные
включаться в цикл биохимических реакций
и регулировать обмен веществ и
энергии.Они
вырабатываются железами внутренней
секреции человека, позвоночных и
некоторых беспозвоночных животных.
Гормоны
могут быть белковой природы(гормон
роста, гормоны поджелудочной железы — инсулин и глюкагон и др.), производными
аминокислот(гормон
щитовидной железы — тироксин, гормоны
надпочечников — адреналин и
норадреналин и др.), липидной природы
(половые
гормоны и др.).
Характерными
особенностями гормонов являются
следующие.
1. Высокая биологическая активность.Даже
незначительные концентрации гормонов
оказывают влияние на клетки, ткани и
органы.
2. Высокая специфичность.Гормонывлияют
только на определенные процессы в
определенных тканях и органах. Они
действуют только на те клетки (так
называемые клетки-мишени), которые имеют
особые рецепторы, способные их
распознавать.
3. Дистанционность действия.Гормоны
с током крови могут переноситься на
значительные расстояния от места их
образования к клеткам-мишеням.
Под контролем
гормонов происходят все этапы
индивидуального развития человека и
животных, а также все процессы
жизнедеятельности. Они обеспечивают
приспособления к изменениям условий
внешней и внутренней среды организма,
регуляцию активности ферментов.
Если определенные
гормоны вырабатываются в недостаточном
количестве или не вырабатываются
вообще в организме, наблюдаются
нарушения развития и обмена веществ
разной степени тяжести. Так же отрицательно
на организм влияет и чрезмерное
образование определенных гормонов. Вы
уже знаете, что при недостаточном
образовании в организме человека гормона
роста развивается карликовость, а при
чрезмерном — гигантизм.
У
растений синтезируются фитогормоны (от
греч. фитон
— растение
и гормао).Как
и гормоны животных, они способны в
малых количествах регулировать и
координировать индивидуальное развитие
и рост растений.
Фитогормоны влияют
практически на все процессы развития
растений: деление и рост клеток,
дифференцирование тканей, формирование
органов, развитие почек, прорастание
семян и т.д. Одни из фитогормонов
стимулируют эти процессы, другие,
наоборот, угнетают.
Например, ауксинактивизирует
деление и растяжение клеток, способствует
формированию, корневой системы. Цитокинины,которых
больше всего в семенах, плодах и в
образовательной ткани, стимулируют
деление клеток. Гиббереллиныусиливают
рост органов растений.
Фитогормонам, как
и гормонам животных, свойственна
дистанционность действия, однако их
специфичность выражена слабее:
различные фитогормоны при определенных
условиях и концентрациях проявляют
подобное действие.
Еще
один вид биологически активных веществ — феромоны (от греч.феро
— несуи хормао— возбуждаю).
Феромоны, как и гормоны, представляют
собой сигнальные молекулы. Однако если
гормоны переносят информацию внутри
организма, то феромоны выделяются во
внешнюю среду и передают химические
сигналы от одного организма к другому.
Они оказывают влияние на поведение и
физиологическое состояние особей своего
вида. Феромоны служат для нахождения
пары, для обозначения своей территории
или для защиты от врагов. Феромоны — чаще всего летучие вещества, которые,
как и гормоны, действуют в малых
концентрациях. Химическая коммуникация
с помощью феромонов наблюдается у
бактерий, протистов и животных.
Алкалоиды—
это органические биологически активные
вещества в основном растительного
происхождения.Большинство
алкалоидов ядовиты для животных и
человека, а некоторые из них оказывают
наркотическое действие (никотин, морфин
и др.). Алкалоиды обнаружены
приблизительно у 2500 видов покрытосеменных
растений. Значение алкалоидов в жизни
растений, по-видимому, заключается в
защите от поедания животными. Некоторые
алкалоиды в малых дозах используются
в медицине в качестве лекарств (атропин,
морфин, кофеин и др.). Алкалоид хинин
применяют при лечении малярии: он
угнетает жизнедеятельность малярийного
плазмодия в эритроцитах человека.
Особая
группа биологически активных веществ —антибиотики(от
греч. анти
— против
и биос
— жизнь)—
биологическиактивные
вещества, вырабатываемые микроорганизмами.Эти
соединения влияют на клетки других
микроорганизмов, тормозя их развитие
или убивая их.
Человек широко
использует антибиотики для лечения
заболеваний, вызванных болезнетворными
бактериями или грибами. Некоторые
антибиотики тормозят рост злокачественных
опухолей, угнетая размножение раковых
клеток.
С помощью биологически
активных соединений организмы оказывают
влияние на организмы своего и других
видов. Так, насекомые с помощью различных
биологически активных веществ способны
привлекать особей противоположного
пола или отпугивать врагов. Растения с
помощью биологически активных веществ
могут угнетать рост других растений.
Взаимовлияние между различными видами
растений человек должен учитывать,
высевая их на одном участке и в
севооборотах.
Пробиотики
– обязательно для современного уровня!
1. Какова структура молекулы АТФ? 2. Какова роль
АТФ в превращении энергии в клетке? 3. Какие связи называются макроэргическими? 4. Какие группы биологически активных
веществ вам известны? 5. Почему при нехватке или отсутствии
определенных витаминов в организме
человека и животных наблюдается нарушение
обмена веществ? 6. Каковы основные свойства гормонов? 5.
Что такое алкалоиды и каково их
значение в жизни человека? 7. Благодаря каким свойствам антибиотики
применяют в медицине? 8. В одну клетку ввели молекулы АТФ, меченные
радиоактивным фосфором Р
по последнему, третьему остатку фосфорной
кислоты, а в другую – молекулы АТФ,
меченные
Р
по первому, ближайшему к рибозе остатку. Через 5 минут в обоих клетках померили
содержание неорганического фосфат-иона,
меченного
Р.
Где оно окажется значительно выше?
Почему?
Почему АТФ так важен?
X
Главная Войти Корзина Информация для заказа Продукция Услуги Конъюгация антител Инструменты Ресурс s О нас Карьера Свяжитесь с нами Дистрибьюторы Условия использования Условия продаж Конфиденциальность
AAT Bioquest
AAT Bioquest
Конъюгация антител
Свяжитесь с нами
Телефон:
1-408-733-1055
Факс: 9003 9
1-408-733-1304
Время работы:
с понедельника по пятницу 8:30 — 17:30 PST (GMT-8)
Адрес:
5775 W Las Positas Blvd. Плезантон, Калифорния, 94588 США
Нажмите здесь, чтобы увидеть всех доступных дистрибьюторов 0002 Ферменты
Клеточный метаболизм
Клеточные процессы
Ответ
Живые клетки не могут хранить значительные количества свободной энергии. Большое количество свободной энергии создаст избыточное тепло в клетке и в конечном итоге разрушит клетку. АТФ (аденозинтрифосфат) действует как эффективное решение. АТФ действует как хранилище энергии клетки. Это позволяет клеткам безопасно хранить энергию в небольших пакетах и высвобождать энергию для использования только по мере необходимости. Другими словами, АТФ служит для закрытия разрыва между реакциями, высвобождающими энергию, такими как расщепление пищи, и действиями, требующими энергии, такими как синтез.
Эта универсальная молекула, названная «энергетической валютой» клетки, хранит энергию, которая высвобождается, когда клетки нуждаются в ней для выполнения основных клеточных функций, включая:
Транспорт макромолекул, таких как липиды и белки, в клетку и из нее
Распространение нервного импульса
Внутриклеточная и внеклеточная передача сигналов
Синтез ДНК и РНК
Сокращение мышц
Обеспечение энергией хромосом и жгутиков для поддержания эффективного функционирования
Фосфорилирование субстрата
АТФ также играет важную роль в поддержании структуры клетки, способствуя сборке компонентов цитоскелета. Все эти ключевые процессы, а также несколько других связанных с ними реакций создают высокую потребность в АТФ.
Дополнительные ресурсы
Модуляторная роль АТФ и аденозина в холинергической нервно-мышечной передаче
Набор для колориметрического анализа АТФ PhosphoWorks™
Связанные вопросы
Являются ли ферменты липидами или белками?
АТФ или АДФ хранят больше энергии?
Как проверить активность ацетилхолинэстеразы?
Сколько молекул АТФ может быть получено при окислении жирных кислот?
Всегда ли энергия выделяется или поглощается во время химической реакции?
Помимо АТФ, новые роли митохондрий | Биохимик
Пропустить пункт назначения
Особенность|
23 августа 2022 г.
Рам Просад Чакрабарти; Навдип С. Чандел
Biochem (Лондон) (2022) 44 (4): 2–8.
https://doi.org/10.1042/bio_2022_119
Разделенный экран
Взгляды
Содержание артикула
Рисунки и таблицы
Видео
Аудио
Дополнительные данные
Экспертная оценка
Нажмите здесь, чтобы открыть pdf в другом окне
PDF для
Делиться
Фейсбук
Твиттер
LinkedIn
Электронная почта
Иконка Цитировать Цитировать
Получить разрешения
Цитирование
Рам Просад Чакрабарти, Навдип С. Чандел; Помимо АТФ, новые роли митохондрий. Biochem (Лондон) 30 августа 2022 г.; 44 (4): 2–8. doi: https://doi.org/10.1042/bio_2022_119
Скачать файл цитирования:
Рис (Зотеро)
Менеджер ссылок
EasyBib
Подставки для книг
Менделей
Бумаги
Конечная примечание
РефВоркс
Бибтекс
поиск панели инструментов
Расширенный поиск
Митохондрии, специальные двухмембранные внутриклеточные компартменты или «органеллы», широко известны как «электростанции клетки», поскольку они генерируют основную часть АТФ, используемого для подпитки клеточных биохимических реакций. Митохондрии также хорошо известны тем, что они генерируют метаболиты для синтеза макромолекул (например, углеводов, белков, липидов и нуклеиновых кислот). В середине 1990-х годов начали появляться новые данные, свидетельствующие о том, что митохондрии, помимо их канонической роли в биоэнергетике и биосинтезе, могут действовать как сигнальные органеллы, что привело к резкому изменению нашего взгляда на роль митохондрий в контроле клеточных функций. В течение следующих двух с половиной десятилетий работы нескольких групп продемонстрировали, как митохондриальная передача сигналов может определять различные физиологические и патофизиологические результаты. В этой статье мы кратко обсудим различные механизмы, с помощью которых митохондрии могут связываться с цитозолем и другими органеллами, чтобы регулировать судьбу и функцию клеток и оказывать паракринные эффекты. Наше молекулярное понимание митохондриальной связи с остальной частью клетки, то есть митохондриальной передачи сигналов, может выявить новые терапевтические стратегии для улучшения здоровья и облегчения заболеваний.
Согласно ведущей эндосимбиотической теории, более 1,45 миллиарда лет назад два прокариота — археон и α-протеобактерия — установили взаимовыгодные или «симбиотические» биологические отношения, чтобы поддерживать потребности друг друга в питании. В конце концов, археи приобрели α-протеобактерии, которые стали первичными митохондриями. Хотя основные механизмы, объясняющие симбиоз между α-протеобактериями и археями, не совсем понятны, текущие данные показывают, что обмен метаболитами между археями-хозяевами и эндосимбионтными α-протеобактериями составляет основу этого симбиоза. Со временем эндосимбионт передал большую часть своей ДНК ядру хозяина, сохранив лишь часть своих генов. На сегодняшний день митохондрии широко распространены среди всех эукариотических линий и продолжают постоянно обмениваться метаболитами с цитозолем.
Рихард Альтманн, немецкий патолог и гистолог, впервые признал повсеместное распространение этих структур, похожих на митохондрии, и ввел для них название «биобласты» в 1890 году. В 1898 году Карл Бенда ввел название «митохондрии» (от греческого « mitos’: нить и ‘chondros’: гранула) для этих структур, имея в виду их появление во время сперматогенеза. Митохондрии представляют собой двухмембранные органеллы с наружной мембраной, обращенной к цитозолю, и внутренней мембраной со складками крист, выступающими в матрикс. Основная часть комплексов электрон-транспортной цепи (ЭТЦ) локализуется в кристах, а ферменты цикла трикарбоновых кислот (ТКА) и митохондриальный геном локализуются в матриксе.
Обширные исследования, проведенные в течение следующих восьми десятилетий, установили две основные функции митохондрий — производство АТФ, энергетического обмена клеток и производство промежуточных продуктов биосинтеза (рис. 1). Цикл лимонной кислоты, также называемый циклом ТСА, происходит в митохондриальном матриксе эукариотических клеток, который может окислять пируват, жирные кислоты и аминокислоты, полученные в результате гликолиза, и генерировать метаболические промежуточные продукты и восстанавливающие эквиваленты (например, НАДН и ФАДН ). 2 ). Эти восстанавливающие эквиваленты подают электроны в ЭТЦ, которая перекачивает протоны через внутреннюю мембрану митохондрий для создания электрохимического градиента, необходимого как для производства АТФ путем окислительного фосфорилирования, так и для переноса белков в митохондрии и из них. Митохондрии поставляют основную часть клеточных АТФ и помогают клеткам поддерживать высокое соотношение АТФ/АДФ, необходимое для термодинамического управления многими биохимическими реакциями, и поэтому широко известны как «электростанции клетки». Кроме того, митохондриальные метаболические промежуточные продукты вступают в различные биосинтетические пути для образования многих важных макромолекул, таких как углеводы, липиды, белки и нуклеотиды. Важно отметить, что митохондрии необходимы для железо-серного кластера и синтеза гема. Наши и другие данные указывают на необходимость биосинтетических функций митохондрий для поддержки пролиферации раковых клеток.
Рисунок 1
УвеличитьСкачать слайд
Митохондрии как биоэнергетические и биосинтетические органеллы. Митохондрии генерируют АТФ посредством окислительного фосфорилирования ( a ) и метаболитов цикла трикарбоновых кислот для поддержки синтеза макромолекул для производства биомассы ( b ).
Рисунок 1
УвеличитьСкачать слайд
Митохондрии как биоэнергетические и биосинтетические органеллы. Митохондрии генерируют АТФ путем окислительного фосфорилирования (9).0310 a ) и метаболиты цикла ТСА для поддержки синтеза макромолекул для производства биомассы ( b ).
Close modal
К 1990 году были открыты многие функциональные свойства митохондрий, и считалось, что в области митохондриальной биохимии осталось не так много исследований. Большое внимание было уделено растущей области митохондриальной генетики. Ключевое наблюдение, которое показало, что митохондрии являются не просто автономными органеллами, но реагируют на цитозольные сигналы, было сделано Розарио Риццуто и его коллегами, которые заметили, что изменения уровня кальция в цитозоле могут вызывать изменения в митохондриальном кальции. Впоследствии «прорывной эксперимент», о котором сообщалось в 1996 лаборатории Xiaodong Wang в Медицинской школе Университета Эмори продемонстрировали, что митохондрии высвобождают цитохром c , ключевой компонент ETC, участвующий в окислительном фосфорилировании, чтобы вызвать особую форму запрограммированной гибели клеток, то есть апоптоз. Таким образом, тот факт, что цитохром c может действовать как сигнальная молекула, помимо его роли в ETC, вызвал новый интерес ученых к изучению других митохондриальных сигнальных механизмов, которые определяют физиологические и патофизиологические результаты. Исследования, проведенные несколькими группами за последние два с половиной десятилетия, твердо установили роль митохондрий как сигнальных органелл (рис. 2). Связь между митохондриями и цитозолем может быть двусторонней: передача сигнала из митохондрий в цитозоль известна как ретроградная передача сигналов, тогда как передача сигнала из цитозоля в митохондрии известна как антероградная передача сигналов. В следующих нескольких разделах мы дадим краткий обзор различных механизмов ретроградной передачи сигналов, с помощью которых митохондрии могут связываться с клеткой и тем самым регулировать физиологические и патофизиологические результаты.
Рисунок 2
Просмотреть в большом размереСкачать слайд
Митохондрии как сигнальные органеллы. Митохондриальные активные формы кислорода (АФК), метаболиты, нуклеиновые кислоты, белки и пептиды, соотношение НАДН/НАД + и морфологическая динамика могут действовать как сигналы для регуляции различных клеточных процессов. AMPK: AMP-активируемая протеинкиназа; HRI: гем-регулируемый ингибитор; GCN2: общий контроль, невыдавливаемый 2; MAVS: митохондриальный противовирусный сигнальный белок; STING: стимулятор генов интерферона; TLR9: толл-подобный рецептор 9; NLRP3: белок, содержащий NOD-, LRR- и пириновый домен 3.
Рисунок 2
Просмотреть большойЗагрузить слайд
Митохондрии как сигнальные органеллы. Митохондриальные активные формы кислорода (АФК), метаболиты, нуклеиновые кислоты, белки и пептиды, соотношение НАДН/НАД + и морфологическая динамика могут действовать как сигналы для регуляции различных клеточных процессов. AMPK: AMP-активируемая протеинкиназа; HRI: гем-регулируемый ингибитор; GCN2: общее управление недепрессируемое 2; MAVS: митохондриальный противовирусный сигнальный белок; STING: стимулятор генов интерферона; TLR9: толл-подобный рецептор 9; NLRP3: белок 3, содержащий NOD-, LRR- и пириновый домен. Сразу после образования супероксиддисмутазы (СОД) восстанавливают супероксид до более стабильного и мембранопроницаемого H 2 O 2 просачивающийся из митохондрий в цитозоль. H 2 O 2 может служить сигнальной молекулой, окисляя определенные серосодержащие аминокислоты, т. е. цистеин и метионин, которые имеют решающее значение для функции, стабильности и субклеточной локализации белка. Вдохновленные обнаружением цитохрома c Xiaodong Wang, мы показали в 1998 году, что высвобождение митохондриальных АФК необходимо для транскрипции генов, вызванной гипоксией. Впоследствии наши текущие работы, а также другие работы продемонстрировали, что генерируемый митохондриями H 2 O 2 может контролировать различные физиологические процессы и заболевания, включая иммунитет, физическую нагрузку, развитие, термогенез, определение кислорода, рак, ишемию-реперфузию, нейродегенерацию и фиброз. Вполне вероятно, что H 2 O 2 вызывает полезные физиологические реакции, тогда как его способность генерировать токсичные гидроксильные радикалы и гидропероксиды липидов вызывает заболевания. Конкретные цистеиновые или метиониновые мишени H 2 O 2 в белках, которые имеют отношение к физиологии или заболеванию, еще полностью не расшифрованы.
Несколько митохондриальных метаболитов могут действовать как сигнальные молекулы, регулируя активность различных цитозольных и ядерных ферментов. Например, метилирование нуклеиновых кислот (т. е. ДНК и РНК), а также метилирование и ацетилирование гистонов, которые контролируют экспрессию генов, могут регулироваться митохондриальными метаболитами. Митохондриальный одноуглеродный метаболизм способствует продукции S -аденозилметионина (SAM), субстрата для гистоновых, ДНК- и РНК-метилтрансфераз. Цитрат метаболита цикла ТСА может экспортироваться в цитозоль и превращаться в ацетил-КоА, субстрат для ацетилирования гистонов. α-Кетоглутарат (α-KG), другой метаболит цикла ТСА, является субстратом для широкого спектра ферментов диоксигеназы, которые регулируют различные клеточные функции. Некоторые примеры α-KG-зависимых диоксигеназ включают деметилазы нуклеиновых кислот и деметилазы гистонов, содержащие домен Jumonji-C, которые удаляют метильные группы из нуклеиновых кислот и гистонов, соответственно, и пролилгидроксилазы, которые регулируют гипоксическую реакцию. Напротив, сукцинат, фумарат и 2-гидроксиглутарат (2-HG) являются конкурентными ингибиторами этих диоксигеназ. В результате клетки могут использовать соотношение α-KG к этим метаболитам для связи между митохондриями и цитозолем/ядром. Как метаболиты находят свои специфические локусы для модуляции экспрессии генов, неизвестно.
Митохондриальный комплекс I ЭТЦ регенерирует НАД + посредством окисления НАДН. Клетки имеют разные механизмы восприятия изменения клеточного, а также митохондриального соотношения НАДН/НАД + . Например, в клетках есть семейство NAD + -зависимых деацилазных ферментов, известных как сиртуины, которые удаляют различные ацильные группы (например, ацетильную, сукцинильную, малонильную, глутарильную или длинноцепочечные ацильные группы) из белков и тем самым регулируют их активность. деятельность. У млекопитающих есть семь сиртуинов (SIRT1–SIRT7), которые локализуются в определенных субклеточных компартментах. SIRT1 и SIRT2 локализуются в цитозоле или ядре; SIRT6 и SIRT7 локализуются в ядре; а SIRT3, SIRT4 и SIRT5 локализуются в митохондриальном матриксе. Увеличение НАДН/НАД 9Соотношение 0340 + может ингибировать ядерный SIRT1, чтобы контролировать окисление жирных кислот и катаболизм аминокислот во время ограничения калорий у мышей. Существует большой интерес к манипулированию соотношением НАДН/НАД + для улучшения здоровья и облегчения заболеваний.
NADH/NAD + также могут напрямую определять судьбу и функцию клеток, регулируя соотношение L-2-HG/α-KG. Гипоксия, кислотность или нарушение митохондриальной ETC снижает регенерацию NAD + и увеличивает NADH/NAD + , который вызывает повышение уровня L-2-HG. Митохондриальные и цитозольные малатдегидрогеназы (MDh2 и MDh3 соответственно) и лактатдегидрогеназы (LDHA и LDHC) могут использовать α-KG в качестве беспорядочного субстрата и NADH в качестве кофермента для производства L-2-HG. Будучи структурно подобным α-KG, L-2-HG может конкурентно ингибировать α-KG-зависимые диоксигеназы, включая PHD2 (домен пролилгидроксилазы-2), деметилазы нуклеиновых кислот и деметилазы гистонов, содержащие домен Jumonji C. PHD2 является ключевым регулятором гипоксического ответа, зависимого от фактора, индуцируемого гипоксией (HIF), а нуклеиновые кислоты и деметилазы гистонов регулируют экспрессию генов. Мы продемонстрировали, что увеличение митохондриального NADH/NAD + вызывает накопление L-2-HG, вызывая нарушение регуляторных Т-клеток (Tregs) в сочетании с широко распространенным аутоиммунитетом из-за гиперметилирования ДНК.
Митохондриальное соотношение NADH/NAD + может также запускать активацию интегрированной реакции на стресс (ISR) у млекопитающих через GCN2 (общий контроль недерепрессивного 2)-зависимого фосфорилирования eIF2α (эукариотический фактор инициации трансляции, вторая субъединица α). Это приводит к ингибированию глобальной трансляции белков, но усиливает трансляцию селективных генов как адаптивный механизм для противодействия метаболическому стрессу. К ним относятся факторы транскрипции (активирующий фактор транскрипции 4 (ATF4) и ATF5), которые увеличивают экспрессию генов, участвующих в метаболизме. Альтернативный механизм включает путь OMA1-DELE1-HRI для запуска ISR. В ответ на митохондриальный стресс, такой как ингибирование АТФ-синтазы или тяжелая деполяризация, ОМА1, протеаза, локализованная во внутренней митохондриальной мембране, расщепляет DELE1 (DAP3-связывающий усилитель клеточной гибели 1), белок, также локализованный во внутренней митохондриальной мембране. Затем расщепленный DELE1 покидает митохондрии и накапливается в цитозоле, где он физически взаимодействует с HRI (ингибитором, регулируемым гемом) и активирует его киназную активность eIF2α, чтобы способствовать трансляции ATF4 ниже фосфорилирования eIF2α. Активация ISR связана с первичными митохондриальными заболеваниями. Однако, является ли это полезной или неадекватной реакцией, до конца не выяснено.
Митохондриальная ДНК (мтДНК) и РНК (мтРНК) могут просачиваться в цитозоль и вызывать иммунный ответ посредством различных сигнальных каскадов. Митохондриальная ДНК с помощью механизмов, которые еще не полностью определены, может просачиваться в цитозоль, где они действуют как молекулярные паттерны, связанные с повреждением (DAMP), чтобы активировать путь cGAS-STING (циклическая GMP-AMP-синтаза-стимулятор генов интерферона), который индуцирует транскрипцию генов, кодирующих интерфероны типа I, такие как интерферон-β (IFNβ), провоспалительные цитокины, такие как интерлейкин 6 (IL-6) и фактор некроза опухоли (TNF). Например, инфекции, вызванные вирусом простого герпеса, могут вызывать высвобождение мтДНК для усиления противовирусного иммунного ответа из-за повышения IFNβ. Поскольку митохондрии являются потомками эндосимбиотических бактерий и имеют кольцевой геном, мтДНК подвергается двунаправленной транскрипции, в результате которой образуются крайне нестабильные длинные митохондриальные dsRNAs (mtdsRNAs), состоящие из кодируемых тяжелой (H) и легкой (L) цепей РНК. Обычно РНК-деградосома быстро разрушает РНК, кодируемую L-цепью. Однако любой дефект образования РНК-деградосом приводит к избыточному накоплению в цитозоле mtdsRNAs, что напоминает маркер dsRNA вирусной репликации и, таким образом, запускает интерфероновый ответ I типа. Следует отметить, что митохондриальный противовирусный сигнальный белок (MAVS), который в основном локализуется на внешней мембране митохондрий, действует как сигнальный узел для интерферон-зависимого иммунного ответа, индуцированного дцРНК. Ключевые оставшиеся без ответа вопросы включают следующее: (1) Как мтДНК или мтРНК высвобождаются в цитозоль, чтобы найти соответствующие им иммунные рецепторы? (2) Почему для оптимального функционирования MAVS требуется локализация внешней мембраны митохондрий?
Митохондрии представляют собой высокодинамичные органеллы, в которых постоянно происходят противоположные процессы слияния (объединения двух митохондрий в одну) и деления (деления одной митохондрии на две), часто называемые «митохондриальной динамикой». Три большие ГТФазы, митофузин 1 (MFN1), митофузин 2 (MFN2) и оптическая атрофия 1 (OPA1), организуют слияние митохондрий в клетках млекопитающих. MFN1 и MFN2 локализуются на внешней мембране митохондрий, а OPA1 локализуется на внутренней мембране митохондрий. Напротив, цитозольный белок, родственный динамину белок 1 (DRP1), перемещается на внешнюю митохондриальную мембрану при активации, чтобы вызвать деление митохондрий млекопитающих. Митохондриальная динамика также включает ремоделирование крист, биогенез и митофагию и связана с апоптозом, а также с функцией стволовых клеток, нейрональных клеток и Т-клеток. Митохондриальная динамика изменяет размер митохондрий, форму, распределение и относительный митохондриальный объем, занимаемый кристами по сравнению с матриксом, и, таким образом, может регулировать производство энергии, синтез макромолекул, Ca 9Передача сигналов 0340 2+ и передача сигналов окислительно-восстановительного потенциала и метаболитов в ответ на клеточный стресс или наличие питательных веществ (рис. 3). Например, когда клетки лишены питательных веществ, они активируют AMPK (AMP-активируемая протеинкиназа), важный датчик клеточной энергии, из-за повышенного уровня AMP по сравнению с АТФ, что способствует делению и вызывает разрушение дефектных митохондрий посредством митофагии.
Рис. 3
Посмотреть большойСкачать слайд
Динамика митохондрий определяет судьбу и функции клеток. Митохондрии морфологически очень динамичны. Они могут передавать сигналы другим частям клеток, изменяя их форму за счет ремоделирования крист, а также противоположных процессов слияния и деления, определяющих судьбу и функции клеток. MFN1/2: митофузин 1/2; OPA1 : атрофия зрительного нерва-1; Рис. 3 Митохондрии морфологически очень динамичны. Они могут передавать сигналы другим частям клеток, изменяя их форму за счет ремоделирования крист, а также противоположных процессов слияния и деления, определяющих судьбу и функции клеток. MFN1/2: митофузин 1/2; OPA1 : атрофия зрительного нерва-1; Drp1: родственный динамину белок 1.
Близкий модальный
Митохондрии физически контактируют с несколькими органеллами, такими как эндоплазматическая сеть (ЭР), лизосомы, аппарат Гольджи, пероксисомы и липидные капли (рис. 4). Контакты митохондрий-ER регулируют передачу сигналов Ca 2+ , передачу сигналов окислительно-восстановительного потенциала, митохондриальную динамику и контроль качества, метаболизм липидов и реакцию развернутых белков. Точно так же контакты митохондрий и лизосом регулируют митохондриальную и лизосомальную динамику и Ca 2+ , и это нарушается мутациями, связанными с болезнью Паркинсона. Хотя есть доказательства физического контакта между митохондриями и пероксисомами, липидными каплями и аппаратом Гольджи, клеточные процессы, регулируемые этими взаимодействиями, еще предстоит полностью расшифровать. Более того, механизм, который специфически связывает эти органеллы, до конца не изучен.
Рисунок 4
УвеличитьЗагрузить слайд
Взаимодействие митохондрий и органелл регулирует различные клеточные процессы. Митохондрии могут взаимодействовать с другими клеточными органеллами, образуя физические контакты для регулирования различных клеточных процессов.
Рисунок 4
УвеличитьЗагрузить слайд
Взаимодействие митохондрий и органелл регулирует различные клеточные процессы. Митохондрии могут взаимодействовать с другими клеточными органеллами, образуя физические контакты для регулирования различных клеточных процессов.
Близкий модальный
Митохондриальный стресс часто побуждает клетки высвобождать растворимые молекулы, такие как метаболиты (например, сукцинат), белки (FGF-21, GDF15) или пептиды (MOTS-c, Humanin), которые часто действуют на другие клетках или тканях паракринным образом, чтобы вызвать системный ответ. Эти сигнальные молекулы также называют «митокинами». Например, сукцинат, метаболит цикла ТСА, который может действовать как внутриклеточная сигнальная молекула, может также действовать как внеклеточная сигнальная молекула для регуляции иммунного ответа, липолиза и восстановления тканей путем связывания с рецептором, связанным с G-белком, SUCNR1 (сукцинатный рецептор). 1), на клетках-мишенях и активации G-белков и нижестоящих эффекторов (рис. 5). Точно так же GDF15 высвобождается из клеток в ответ на митохондриальный стресс, связывается с рецепторами GFRAL (GDNF (нейротрофический фактор, полученный из глиальной клеточной линии) семейство α-подобных рецепторов) на клетках-мишенях и активирует нижестоящие эффекторы для регулирования массы тела, потребления пищи, глюкозы. метаболизм и иммунный ответ (рис. 5).
Рисунок 5
УвеличитьЗагрузить слайд
Митохондриальная сигнализация может систематически регулировать физиологические и патологические процессы. Митохондриальный стресс может побуждать клетку высвобождать сигнальные молекулы, называемые «митокинами» (например, сукцинат и GDF15), которые часто действуют на другие клетки паракринным образом, определяя физиологические и патологические результаты. ISR: интегрированная реакция на стресс ; ATF4: активирующий транскрипционный фактор 4; ГДФ15: фактор дифференциации роста 15; SUCNR1: сукцинатный рецептор 1; GFRAL: GDNF (нейротрофический фактор, полученный из глиальной клеточной линии) семейство α-подобных рецепторов.
Рисунок 5
УвеличитьЗагрузить слайд
Митохондриальная передача сигналов может систематически регулировать физиологические и патологические процессы. Митохондриальный стресс может побуждать клетку высвобождать сигнальные молекулы, называемые «митокинами» (например, сукцинат и GDF15), которые часто действуют на другие клетки паракринным образом, определяя физиологические и патологические результаты. ISR: комплексная реакция на стресс; ATF4: активирующий транскрипционный фактор 4; GDF15: фактор дифференциации роста 15; SUCNR1: сукцинатный рецептор 1; GFRAL: GDNF (нейротрофический фактор, полученный из глиальной клеточной линии) семейство α-подобных рецепторов.
Close modal
В прошлом веке митохондрии стали биоэнергетическими и биосинтетическими органеллами, поддерживающими выживание клеток и производство биомассы соответственно. Многие учебники показывают судьбу молекул углерода через промежуточный метаболизм. С начала этого века большой интерес к митохондриям был связан с их ролью сигнальных органелл в контроле физиологии и болезней. Хотя концептуальная основа митохондрий как сигнальных органелл уже установлена, существует столь необходимое понимание детальных механизмов. Есть надежда, что появятся новые мишени для лечения заболеваний, связанных с митохондриями, таких как болезнь Паркинсона, первичные митохондриальные заболевания и заболевания, связанные с воспалением, по мере выяснения молекулярных деталей, лежащих в основе митохондрий как парадигмы сигнальных органелл.
НСК получает финансовую поддержку от NIH (5R35ACA197532, 5R01AI148190, 5P01AG049665 и 1P01HL154998-01A15670) и Фонда Майкла Дж. Фокса для исследований болезни Паркинсона; Р.П.К. поддерживается преддокторской стипендией Кугелла отделения пульмонологии и интенсивной терапии Северо-Западного университета. Все фигуры созданы с помощью BioRender.com.
ДеБерардинис Р.Дж. и Чандел Н.С. (2020). Нам нужно поговорить об эффекте Варбурга. Нац. Метаб ., 2 , 127–129. DOI: 10.1038/s42255-020-0172-2
Джакомелло М. , Пякурель А., Глыцоу К. и Скоррано Л. (2020). Клеточная биология динамики митохондриальной мембраны. Нац. Преподобный Мол. Cell Biol ., 21 , 204–224. DOI: 10.1038/s41580-020-0210-7
Райли, Дж. С., и Тейт, С. В. (2020). Митохондриальная ДНК при воспалении и иммунитете. EMBO Реп ., 21 , е49799. DOI: 10.15252/embr.201949799
Сиес Х., Белоусов В.В., Чандел Н.С., Дэвис М.Дж., Джонс Д.П., Манн Г.Е., Джонс Д.П., Манн Г.Е., Мерфи М.П., Яма мото, М. и Уинтерборн, К. (2022). Определение роли конкретных активных форм кислорода (АФК) в клеточной биологии и физиологии. Нац. Преподобный Мол. Cell Biol ., 499–515. DOI: 10.1038/s41580-022-00456-z
Скоррано, Л., Де Маттеис, М.А., Эмр, С., Джордано, Ф., Хайноцки, Г., Корнманн, Б., Лакнер, Л.Л., Левин , Т.П., Пеллегрини, Л., Райниш, К., и Шульдинер, М. (2019). Собираясь вместе, чтобы определить места контакта с мембраной. Нац. Comm ., 10 , 1287. DOI: 10.1038/s41467-019-09253-3
Кацюба, Э., Романи, М., Хофер, Д., и Ауверкс, Дж. (2020). НАД + Гомеостаз в норме и при болезни. Нац. Метаб ., 2 , 9–31. DOI: 10.1038/s42255-019-0161-5
Рам Просад Чакрабарти — кандидат медицинских наук в лаборатории профессора Навдипа Чанделя в Медицинской школе Файнберга Северо-Западного университета. Его исследования сосредоточены на понимании того, как митохондриальная передача сигналов регулирует физиологические и патологические результаты. Он получил степень бакалавра (с отличием) и магистра микробиологии в Университете Дакки, Бангладеш. Твиттер: @RPC_3312
Навдип С. Чандел — заслуженный профессор медицины, биохимии и молекулярной генетики Дэвида В. Кугелла в Северо-Западном университете. Он получил степень бакалавра математики (1991 г.) и докторскую степень в области клеточной физиологии в Чикагском университете (1993–1997 гг.

 трофічну;
B.захисну;
C.ендокринну;
D.імунну;
E. кровотворну
32.Що таке нейрула?:
A.нервова трубка;
B.зародок на стадії утворення нервової трубки;
C.нервові гребені;
D.гангліозні пластинки;
E. порожнина нервової трубки
33.З яких клітин складається бластула
A. Трофобластів.
B. Фолікулярних.
C. Децидуальних.
D.Бластомерів
E. Амніотичних.
35.З якої складової частини бластоцисти утворюється хоріон?
A. Трофобласта
B. Бластоцелі.
C. Амніобласта.
D. Ембріобласта.
E. Бластомерів.
37.Основна біологічна роль яйцеклітини в процесі запліднення:
A. Зустріч із сперматозоїдом.
B. Доповнення диплоїдного набору хромосом, трофічна.
C. Визначення статі зародка
D. Забезпечення моноспермії.
E. Захисна.
38.Основні структурні компоненти головки сперматозоїда:
A. Диплоїдне ядро і акросома.
B. Гаплоїдне ядро і мітохондрії.
C. Диплоїдне ядро і мітохондрії.
D. Гаплоїдне ядро і акросома.
E. Гаплоїдне ядро, акросома і мітохондрії.
39. Перебуваючи на стадії дроблення зародок побудований з клітин:
A. Бластомерів.
B. Трофобластів.
трофічну;
B.захисну;
C.ендокринну;
D.імунну;
E. кровотворну
32.Що таке нейрула?:
A.нервова трубка;
B.зародок на стадії утворення нервової трубки;
C.нервові гребені;
D.гангліозні пластинки;
E. порожнина нервової трубки
33.З яких клітин складається бластула
A. Трофобластів.
B. Фолікулярних.
C. Децидуальних.
D.Бластомерів
E. Амніотичних.
35.З якої складової частини бластоцисти утворюється хоріон?
A. Трофобласта
B. Бластоцелі.
C. Амніобласта.
D. Ембріобласта.
E. Бластомерів.
37.Основна біологічна роль яйцеклітини в процесі запліднення:
A. Зустріч із сперматозоїдом.
B. Доповнення диплоїдного набору хромосом, трофічна.
C. Визначення статі зародка
D. Забезпечення моноспермії.
E. Захисна.
38.Основні структурні компоненти головки сперматозоїда:
A. Диплоїдне ядро і акросома.
B. Гаплоїдне ядро і мітохондрії.
C. Диплоїдне ядро і мітохондрії.
D. Гаплоїдне ядро і акросома.
E. Гаплоїдне ядро, акросома і мітохондрії.
39. Перебуваючи на стадії дроблення зародок побудований з клітин:
A. Бластомерів.
B. Трофобластів.
 Нуклеотиды являются структурной основой
для синтеза целого ряда важных для
жизнедеятельности органических веществ.
Наиболее широко распространенными
среди них являются макроэргические
соединения (вещества, содержащие богатые
энергией, или макроэргические связи).
К таким веществам в первую очередь
относится аденозинтрифосфорная
кислота (АТФ). АТФ – универсальный
хранитель и переносчик энергии в клетках
всех организмов. Практически все идущие
с затратой энергии биохимические
процессы в клетках организмов в качестве
ее источника используют АТФ. С
использованием энергии АТФ в клетке
синтезируют новые молекулы белков,
углеводов, жиров, осуществляется
активный транспорт веществ, биение
жгутиков и ресничек, поддерживается
постоянная температура тела теплокровных
организмов и т.д.
Нуклеотиды являются структурной основой
для синтеза целого ряда важных для
жизнедеятельности органических веществ.
Наиболее широко распространенными
среди них являются макроэргические
соединения (вещества, содержащие богатые
энергией, или макроэргические связи).
К таким веществам в первую очередь
относится аденозинтрифосфорная
кислота (АТФ). АТФ – универсальный
хранитель и переносчик энергии в клетках
всех организмов. Практически все идущие
с затратой энергии биохимические
процессы в клетках организмов в качестве
ее источника используют АТФ. С
использованием энергии АТФ в клетке
синтезируют новые молекулы белков,
углеводов, жиров, осуществляется
активный транспорт веществ, биение
жгутиков и ресничек, поддерживается
постоянная температура тела теплокровных
организмов и т.д. ).
).
 Они принимают участие в
обмене веществ и превращении энергии,
преимущественно как компоненты сложных
ферментов.
Они принимают участие в
обмене веществ и превращении энергии,
преимущественно как компоненты сложных
ферментов.
 гормао
— двигаю,
побуждаю) — органические вещества, способные
включаться в цикл биохимических реакций
и регулировать обмен веществ и
энергии. Они
вырабатываются железами внутренней
секреции человека, позвоночных и
некоторых беспозвоночных животных.
гормао
— двигаю,
побуждаю) — органические вещества, способные
включаться в цикл биохимических реакций
и регулировать обмен веществ и
энергии. Они
вырабатываются железами внутренней
секреции человека, позвоночных и
некоторых беспозвоночных животных.
 Как
и гормоны животных, они способны в
малых количествах регулировать и
координировать индивидуальное развитие
и рост растений.
Как
и гормоны животных, они способны в
малых количествах регулировать и
координировать индивидуальное развитие
и рост растений. феро
— несу и хормао — возбуждаю).
Феромоны, как и гормоны, представляют
собой сигнальные молекулы. Однако если
гормоны переносят информацию внутри
организма, то феромоны выделяются во
внешнюю среду и передают химические
сигналы от одного организма к другому.
Они оказывают влияние на поведение и
физиологическое состояние особей своего
вида. Феромоны служат для нахождения
пары, для обозначения своей территории
или для защиты от врагов. Феромоны — чаще всего летучие вещества, которые,
как и гормоны, действуют в малых
концентрациях. Химическая коммуникация
с помощью феромонов наблюдается у
бактерий, протистов и животных.
феро
— несу и хормао — возбуждаю).
Феромоны, как и гормоны, представляют
собой сигнальные молекулы. Однако если
гормоны переносят информацию внутри
организма, то феромоны выделяются во
внешнюю среду и передают химические
сигналы от одного организма к другому.
Они оказывают влияние на поведение и
физиологическое состояние особей своего
вида. Феромоны служат для нахождения
пары, для обозначения своей территории
или для защиты от врагов. Феромоны — чаще всего летучие вещества, которые,
как и гормоны, действуют в малых
концентрациях. Химическая коммуникация
с помощью феромонов наблюдается у
бактерий, протистов и животных. Значение алкалоидов в жизни
растений, по-видимому, заключается в
защите от поедания животными. Некоторые
алкалоиды в малых дозах используются
в медицине в качестве лекарств (атропин,
морфин, кофеин и др.). Алкалоид хинин
применяют при лечении малярии: он
угнетает жизнедеятельность малярийного
плазмодия в эритроцитах человека.
Значение алкалоидов в жизни
растений, по-видимому, заключается в
защите от поедания животными. Некоторые
алкалоиды в малых дозах используются
в медицине в качестве лекарств (атропин,
морфин, кофеин и др.). Алкалоид хинин
применяют при лечении малярии: он
угнетает жизнедеятельность малярийного
плазмодия в эритроцитах человека. Так, насекомые с помощью различных
биологически активных веществ способны
привлекать особей противоположного
пола или отпугивать врагов. Растения с
помощью биологически активных веществ
могут угнетать рост других растений.
Взаимовлияние между различными видами
растений человек должен учитывать,
высевая их на одном участке и в
севооборотах.
Так, насекомые с помощью различных
биологически активных веществ способны
привлекать особей противоположного
пола или отпугивать врагов. Растения с
помощью биологически активных веществ
могут угнетать рост других растений.
Взаимовлияние между различными видами
растений человек должен учитывать,
высевая их на одном участке и в
севооборотах. Через 5 минут в обоих клетках померили
содержание неорганического фосфат-иона,
меченного
Р.
Где оно окажется значительно выше?
Почему?
Через 5 минут в обоих клетках померили
содержание неорганического фосфат-иона,
меченного
Р.
Где оно окажется значительно выше?
Почему? com
com Это позволяет клеткам безопасно хранить энергию в небольших пакетах и высвобождать энергию для использования только по мере необходимости. Другими словами, АТФ служит для закрытия разрыва между реакциями, высвобождающими энергию, такими как расщепление пищи, и действиями, требующими энергии, такими как синтез.
Это позволяет клеткам безопасно хранить энергию в небольших пакетах и высвобождать энергию для использования только по мере необходимости. Другими словами, АТФ служит для закрытия разрыва между реакциями, высвобождающими энергию, такими как расщепление пищи, и действиями, требующими энергии, такими как синтез. Все эти ключевые процессы, а также несколько других связанных с ними реакций создают высокую потребность в АТФ.
Все эти ключевые процессы, а также несколько других связанных с ними реакций создают высокую потребность в АТФ. Чандел; Помимо АТФ, новые роли митохондрий. Biochem (Лондон) 30 августа 2022 г.; 44 (4): 2–8. doi: https://doi.org/10.1042/bio_2022_119
Чандел; Помимо АТФ, новые роли митохондрий. Biochem (Лондон) 30 августа 2022 г.; 44 (4): 2–8. doi: https://doi.org/10.1042/bio_2022_119 В течение следующих двух с половиной десятилетий работы нескольких групп продемонстрировали, как митохондриальная передача сигналов может определять различные физиологические и патофизиологические результаты. В этой статье мы кратко обсудим различные механизмы, с помощью которых митохондрии могут связываться с цитозолем и другими органеллами, чтобы регулировать судьбу и функцию клеток и оказывать паракринные эффекты. Наше молекулярное понимание митохондриальной связи с остальной частью клетки, то есть митохондриальной передачи сигналов, может выявить новые терапевтические стратегии для улучшения здоровья и облегчения заболеваний.
В течение следующих двух с половиной десятилетий работы нескольких групп продемонстрировали, как митохондриальная передача сигналов может определять различные физиологические и патофизиологические результаты. В этой статье мы кратко обсудим различные механизмы, с помощью которых митохондрии могут связываться с цитозолем и другими органеллами, чтобы регулировать судьбу и функцию клеток и оказывать паракринные эффекты. Наше молекулярное понимание митохондриальной связи с остальной частью клетки, то есть митохондриальной передачи сигналов, может выявить новые терапевтические стратегии для улучшения здоровья и облегчения заболеваний. Со временем эндосимбионт передал большую часть своей ДНК ядру хозяина, сохранив лишь часть своих генов. На сегодняшний день митохондрии широко распространены среди всех эукариотических линий и продолжают постоянно обмениваться метаболитами с цитозолем.
Со временем эндосимбионт передал большую часть своей ДНК ядру хозяина, сохранив лишь часть своих генов. На сегодняшний день митохондрии широко распространены среди всех эукариотических линий и продолжают постоянно обмениваться метаболитами с цитозолем. 1). Цикл лимонной кислоты, также называемый циклом ТСА, происходит в митохондриальном матриксе эукариотических клеток, который может окислять пируват, жирные кислоты и аминокислоты, полученные в результате гликолиза, и генерировать метаболические промежуточные продукты и восстанавливающие эквиваленты (например, НАДН и ФАДН ). 2 ). Эти восстанавливающие эквиваленты подают электроны в ЭТЦ, которая перекачивает протоны через внутреннюю мембрану митохондрий для создания электрохимического градиента, необходимого как для производства АТФ путем окислительного фосфорилирования, так и для переноса белков в митохондрии и из них. Митохондрии поставляют основную часть клеточных АТФ и помогают клеткам поддерживать высокое соотношение АТФ/АДФ, необходимое для термодинамического управления многими биохимическими реакциями, и поэтому широко известны как «электростанции клетки». Кроме того, митохондриальные метаболические промежуточные продукты вступают в различные биосинтетические пути для образования многих важных макромолекул, таких как углеводы, липиды, белки и нуклеотиды.
1). Цикл лимонной кислоты, также называемый циклом ТСА, происходит в митохондриальном матриксе эукариотических клеток, который может окислять пируват, жирные кислоты и аминокислоты, полученные в результате гликолиза, и генерировать метаболические промежуточные продукты и восстанавливающие эквиваленты (например, НАДН и ФАДН ). 2 ). Эти восстанавливающие эквиваленты подают электроны в ЭТЦ, которая перекачивает протоны через внутреннюю мембрану митохондрий для создания электрохимического градиента, необходимого как для производства АТФ путем окислительного фосфорилирования, так и для переноса белков в митохондрии и из них. Митохондрии поставляют основную часть клеточных АТФ и помогают клеткам поддерживать высокое соотношение АТФ/АДФ, необходимое для термодинамического управления многими биохимическими реакциями, и поэтому широко известны как «электростанции клетки». Кроме того, митохондриальные метаболические промежуточные продукты вступают в различные биосинтетические пути для образования многих важных макромолекул, таких как углеводы, липиды, белки и нуклеотиды. Важно отметить, что митохондрии необходимы для железо-серного кластера и синтеза гема. Наши и другие данные указывают на необходимость биосинтетических функций митохондрий для поддержки пролиферации раковых клеток.
Важно отметить, что митохондрии необходимы для железо-серного кластера и синтеза гема. Наши и другие данные указывают на необходимость биосинтетических функций митохондрий для поддержки пролиферации раковых клеток. Большое внимание было уделено растущей области митохондриальной генетики. Ключевое наблюдение, которое показало, что митохондрии являются не просто автономными органеллами, но реагируют на цитозольные сигналы, было сделано Розарио Риццуто и его коллегами, которые заметили, что изменения уровня кальция в цитозоле могут вызывать изменения в митохондриальном кальции. Впоследствии «прорывной эксперимент», о котором сообщалось в 1996 лаборатории Xiaodong Wang в Медицинской школе Университета Эмори продемонстрировали, что митохондрии высвобождают цитохром c , ключевой компонент ETC, участвующий в окислительном фосфорилировании, чтобы вызвать особую форму запрограммированной гибели клеток, то есть апоптоз. Таким образом, тот факт, что цитохром c может действовать как сигнальная молекула, помимо его роли в ETC, вызвал новый интерес ученых к изучению других митохондриальных сигнальных механизмов, которые определяют физиологические и патофизиологические результаты. Исследования, проведенные несколькими группами за последние два с половиной десятилетия, твердо установили роль митохондрий как сигнальных органелл (рис.
Большое внимание было уделено растущей области митохондриальной генетики. Ключевое наблюдение, которое показало, что митохондрии являются не просто автономными органеллами, но реагируют на цитозольные сигналы, было сделано Розарио Риццуто и его коллегами, которые заметили, что изменения уровня кальция в цитозоле могут вызывать изменения в митохондриальном кальции. Впоследствии «прорывной эксперимент», о котором сообщалось в 1996 лаборатории Xiaodong Wang в Медицинской школе Университета Эмори продемонстрировали, что митохондрии высвобождают цитохром c , ключевой компонент ETC, участвующий в окислительном фосфорилировании, чтобы вызвать особую форму запрограммированной гибели клеток, то есть апоптоз. Таким образом, тот факт, что цитохром c может действовать как сигнальная молекула, помимо его роли в ETC, вызвал новый интерес ученых к изучению других митохондриальных сигнальных механизмов, которые определяют физиологические и патофизиологические результаты. Исследования, проведенные несколькими группами за последние два с половиной десятилетия, твердо установили роль митохондрий как сигнальных органелл (рис. 2). Связь между митохондриями и цитозолем может быть двусторонней: передача сигнала из митохондрий в цитозоль известна как ретроградная передача сигналов, тогда как передача сигнала из цитозоля в митохондрии известна как антероградная передача сигналов. В следующих нескольких разделах мы дадим краткий обзор различных механизмов ретроградной передачи сигналов, с помощью которых митохондрии могут связываться с клеткой и тем самым регулировать физиологические и патофизиологические результаты.
2). Связь между митохондриями и цитозолем может быть двусторонней: передача сигнала из митохондрий в цитозоль известна как ретроградная передача сигналов, тогда как передача сигнала из цитозоля в митохондрии известна как антероградная передача сигналов. В следующих нескольких разделах мы дадим краткий обзор различных механизмов ретроградной передачи сигналов, с помощью которых митохондрии могут связываться с клеткой и тем самым регулировать физиологические и патофизиологические результаты.
 е. цистеин и метионин, которые имеют решающее значение для функции, стабильности и субклеточной локализации белка. Вдохновленные обнаружением цитохрома c Xiaodong Wang, мы показали в 1998 году, что высвобождение митохондриальных АФК необходимо для транскрипции генов, вызванной гипоксией. Впоследствии наши текущие работы, а также другие работы продемонстрировали, что генерируемый митохондриями H 2 O 2 может контролировать различные физиологические процессы и заболевания, включая иммунитет, физическую нагрузку, развитие, термогенез, определение кислорода, рак, ишемию-реперфузию, нейродегенерацию и фиброз. Вполне вероятно, что H 2 O 2 вызывает полезные физиологические реакции, тогда как его способность генерировать токсичные гидроксильные радикалы и гидропероксиды липидов вызывает заболевания. Конкретные цистеиновые или метиониновые мишени H 2 O 2 в белках, которые имеют отношение к физиологии или заболеванию, еще полностью не расшифрованы.
е. цистеин и метионин, которые имеют решающее значение для функции, стабильности и субклеточной локализации белка. Вдохновленные обнаружением цитохрома c Xiaodong Wang, мы показали в 1998 году, что высвобождение митохондриальных АФК необходимо для транскрипции генов, вызванной гипоксией. Впоследствии наши текущие работы, а также другие работы продемонстрировали, что генерируемый митохондриями H 2 O 2 может контролировать различные физиологические процессы и заболевания, включая иммунитет, физическую нагрузку, развитие, термогенез, определение кислорода, рак, ишемию-реперфузию, нейродегенерацию и фиброз. Вполне вероятно, что H 2 O 2 вызывает полезные физиологические реакции, тогда как его способность генерировать токсичные гидроксильные радикалы и гидропероксиды липидов вызывает заболевания. Конкретные цистеиновые или метиониновые мишени H 2 O 2 в белках, которые имеют отношение к физиологии или заболеванию, еще полностью не расшифрованы.
 Напротив, сукцинат, фумарат и 2-гидроксиглутарат (2-HG) являются конкурентными ингибиторами этих диоксигеназ. В результате клетки могут использовать соотношение α-KG к этим метаболитам для связи между митохондриями и цитозолем/ядром. Как метаболиты находят свои специфические локусы для модуляции экспрессии генов, неизвестно.
Напротив, сукцинат, фумарат и 2-гидроксиглутарат (2-HG) являются конкурентными ингибиторами этих диоксигеназ. В результате клетки могут использовать соотношение α-KG к этим метаболитам для связи между митохондриями и цитозолем/ядром. Как метаболиты находят свои специфические локусы для модуляции экспрессии генов, неизвестно. Увеличение НАДН/НАД 9Соотношение 0340 + может ингибировать ядерный SIRT1, чтобы контролировать окисление жирных кислот и катаболизм аминокислот во время ограничения калорий у мышей. Существует большой интерес к манипулированию соотношением НАДН/НАД + для улучшения здоровья и облегчения заболеваний.
Увеличение НАДН/НАД 9Соотношение 0340 + может ингибировать ядерный SIRT1, чтобы контролировать окисление жирных кислот и катаболизм аминокислот во время ограничения калорий у мышей. Существует большой интерес к манипулированию соотношением НАДН/НАД + для улучшения здоровья и облегчения заболеваний. PHD2 является ключевым регулятором гипоксического ответа, зависимого от фактора, индуцируемого гипоксией (HIF), а нуклеиновые кислоты и деметилазы гистонов регулируют экспрессию генов. Мы продемонстрировали, что увеличение митохондриального NADH/NAD + вызывает накопление L-2-HG, вызывая нарушение регуляторных Т-клеток (Tregs) в сочетании с широко распространенным аутоиммунитетом из-за гиперметилирования ДНК.
PHD2 является ключевым регулятором гипоксического ответа, зависимого от фактора, индуцируемого гипоксией (HIF), а нуклеиновые кислоты и деметилазы гистонов регулируют экспрессию генов. Мы продемонстрировали, что увеличение митохондриального NADH/NAD + вызывает накопление L-2-HG, вызывая нарушение регуляторных Т-клеток (Tregs) в сочетании с широко распространенным аутоиммунитетом из-за гиперметилирования ДНК. Альтернативный механизм включает путь OMA1-DELE1-HRI для запуска ISR. В ответ на митохондриальный стресс, такой как ингибирование АТФ-синтазы или тяжелая деполяризация, ОМА1, протеаза, локализованная во внутренней митохондриальной мембране, расщепляет DELE1 (DAP3-связывающий усилитель клеточной гибели 1), белок, также локализованный во внутренней митохондриальной мембране. Затем расщепленный DELE1 покидает митохондрии и накапливается в цитозоле, где он физически взаимодействует с HRI (ингибитором, регулируемым гемом) и активирует его киназную активность eIF2α, чтобы способствовать трансляции ATF4 ниже фосфорилирования eIF2α. Активация ISR связана с первичными митохондриальными заболеваниями. Однако, является ли это полезной или неадекватной реакцией, до конца не выяснено.
Альтернативный механизм включает путь OMA1-DELE1-HRI для запуска ISR. В ответ на митохондриальный стресс, такой как ингибирование АТФ-синтазы или тяжелая деполяризация, ОМА1, протеаза, локализованная во внутренней митохондриальной мембране, расщепляет DELE1 (DAP3-связывающий усилитель клеточной гибели 1), белок, также локализованный во внутренней митохондриальной мембране. Затем расщепленный DELE1 покидает митохондрии и накапливается в цитозоле, где он физически взаимодействует с HRI (ингибитором, регулируемым гемом) и активирует его киназную активность eIF2α, чтобы способствовать трансляции ATF4 ниже фосфорилирования eIF2α. Активация ISR связана с первичными митохондриальными заболеваниями. Однако, является ли это полезной или неадекватной реакцией, до конца не выяснено. Например, инфекции, вызванные вирусом простого герпеса, могут вызывать высвобождение мтДНК для усиления противовирусного иммунного ответа из-за повышения IFNβ. Поскольку митохондрии являются потомками эндосимбиотических бактерий и имеют кольцевой геном, мтДНК подвергается двунаправленной транскрипции, в результате которой образуются крайне нестабильные длинные митохондриальные dsRNAs (mtdsRNAs), состоящие из кодируемых тяжелой (H) и легкой (L) цепей РНК. Обычно РНК-деградосома быстро разрушает РНК, кодируемую L-цепью. Однако любой дефект образования РНК-деградосом приводит к избыточному накоплению в цитозоле mtdsRNAs, что напоминает маркер dsRNA вирусной репликации и, таким образом, запускает интерфероновый ответ I типа. Следует отметить, что митохондриальный противовирусный сигнальный белок (MAVS), который в основном локализуется на внешней мембране митохондрий, действует как сигнальный узел для интерферон-зависимого иммунного ответа, индуцированного дцРНК. Ключевые оставшиеся без ответа вопросы включают следующее: (1) Как мтДНК или мтРНК высвобождаются в цитозоль, чтобы найти соответствующие им иммунные рецепторы? (2) Почему для оптимального функционирования MAVS требуется локализация внешней мембраны митохондрий?
Например, инфекции, вызванные вирусом простого герпеса, могут вызывать высвобождение мтДНК для усиления противовирусного иммунного ответа из-за повышения IFNβ. Поскольку митохондрии являются потомками эндосимбиотических бактерий и имеют кольцевой геном, мтДНК подвергается двунаправленной транскрипции, в результате которой образуются крайне нестабильные длинные митохондриальные dsRNAs (mtdsRNAs), состоящие из кодируемых тяжелой (H) и легкой (L) цепей РНК. Обычно РНК-деградосома быстро разрушает РНК, кодируемую L-цепью. Однако любой дефект образования РНК-деградосом приводит к избыточному накоплению в цитозоле mtdsRNAs, что напоминает маркер dsRNA вирусной репликации и, таким образом, запускает интерфероновый ответ I типа. Следует отметить, что митохондриальный противовирусный сигнальный белок (MAVS), который в основном локализуется на внешней мембране митохондрий, действует как сигнальный узел для интерферон-зависимого иммунного ответа, индуцированного дцРНК. Ключевые оставшиеся без ответа вопросы включают следующее: (1) Как мтДНК или мтРНК высвобождаются в цитозоль, чтобы найти соответствующие им иммунные рецепторы? (2) Почему для оптимального функционирования MAVS требуется локализация внешней мембраны митохондрий? Три большие ГТФазы, митофузин 1 (MFN1), митофузин 2 (MFN2) и оптическая атрофия 1 (OPA1), организуют слияние митохондрий в клетках млекопитающих. MFN1 и MFN2 локализуются на внешней мембране митохондрий, а OPA1 локализуется на внутренней мембране митохондрий. Напротив, цитозольный белок, родственный динамину белок 1 (DRP1), перемещается на внешнюю митохондриальную мембрану при активации, чтобы вызвать деление митохондрий млекопитающих. Митохондриальная динамика также включает ремоделирование крист, биогенез и митофагию и связана с апоптозом, а также с функцией стволовых клеток, нейрональных клеток и Т-клеток. Митохондриальная динамика изменяет размер митохондрий, форму, распределение и относительный митохондриальный объем, занимаемый кристами по сравнению с матриксом, и, таким образом, может регулировать производство энергии, синтез макромолекул, Ca 9Передача сигналов 0340 2+ и передача сигналов окислительно-восстановительного потенциала и метаболитов в ответ на клеточный стресс или наличие питательных веществ (рис.
Три большие ГТФазы, митофузин 1 (MFN1), митофузин 2 (MFN2) и оптическая атрофия 1 (OPA1), организуют слияние митохондрий в клетках млекопитающих. MFN1 и MFN2 локализуются на внешней мембране митохондрий, а OPA1 локализуется на внутренней мембране митохондрий. Напротив, цитозольный белок, родственный динамину белок 1 (DRP1), перемещается на внешнюю митохондриальную мембрану при активации, чтобы вызвать деление митохондрий млекопитающих. Митохондриальная динамика также включает ремоделирование крист, биогенез и митофагию и связана с апоптозом, а также с функцией стволовых клеток, нейрональных клеток и Т-клеток. Митохондриальная динамика изменяет размер митохондрий, форму, распределение и относительный митохондриальный объем, занимаемый кристами по сравнению с матриксом, и, таким образом, может регулировать производство энергии, синтез макромолекул, Ca 9Передача сигналов 0340 2+ и передача сигналов окислительно-восстановительного потенциала и метаболитов в ответ на клеточный стресс или наличие питательных веществ (рис. 3). Например, когда клетки лишены питательных веществ, они активируют AMPK (AMP-активируемая протеинкиназа), важный датчик клеточной энергии, из-за повышенного уровня AMP по сравнению с АТФ, что способствует делению и вызывает разрушение дефектных митохондрий посредством митофагии.
3). Например, когда клетки лишены питательных веществ, они активируют AMPK (AMP-активируемая протеинкиназа), важный датчик клеточной энергии, из-за повышенного уровня AMP по сравнению с АТФ, что способствует делению и вызывает разрушение дефектных митохондрий посредством митофагии.
 Митохондрии могут взаимодействовать с другими клеточными органеллами, образуя физические контакты для регулирования различных клеточных процессов.
Митохондрии могут взаимодействовать с другими клеточными органеллами, образуя физические контакты для регулирования различных клеточных процессов. 1), на клетках-мишенях и активации G-белков и нижестоящих эффекторов (рис. 5). Точно так же GDF15 высвобождается из клеток в ответ на митохондриальный стресс, связывается с рецепторами GFRAL (GDNF (нейротрофический фактор, полученный из глиальной клеточной линии) семейство α-подобных рецепторов) на клетках-мишенях и активирует нижестоящие эффекторы для регулирования массы тела, потребления пищи, глюкозы. метаболизм и иммунный ответ (рис. 5).
1), на клетках-мишенях и активации G-белков и нижестоящих эффекторов (рис. 5). Точно так же GDF15 высвобождается из клеток в ответ на митохондриальный стресс, связывается с рецепторами GFRAL (GDNF (нейротрофический фактор, полученный из глиальной клеточной линии) семейство α-подобных рецепторов) на клетках-мишенях и активирует нижестоящие эффекторы для регулирования массы тела, потребления пищи, глюкозы. метаболизм и иммунный ответ (рис. 5).
 С начала этого века большой интерес к митохондриям был связан с их ролью сигнальных органелл в контроле физиологии и болезней. Хотя концептуальная основа митохондрий как сигнальных органелл уже установлена, существует столь необходимое понимание детальных механизмов. Есть надежда, что появятся новые мишени для лечения заболеваний, связанных с митохондриями, таких как болезнь Паркинсона, первичные митохондриальные заболевания и заболевания, связанные с воспалением, по мере выяснения молекулярных деталей, лежащих в основе митохондрий как парадигмы сигнальных органелл.
С начала этого века большой интерес к митохондриям был связан с их ролью сигнальных органелл в контроле физиологии и болезней. Хотя концептуальная основа митохондрий как сигнальных органелл уже установлена, существует столь необходимое понимание детальных механизмов. Есть надежда, что появятся новые мишени для лечения заболеваний, связанных с митохондриями, таких как болезнь Паркинсона, первичные митохондриальные заболевания и заболевания, связанные с воспалением, по мере выяснения молекулярных деталей, лежащих в основе митохондрий как парадигмы сигнальных органелл. Нац. Метаб ., 2 , 127–129. DOI: 10.1038/s42255-020-0172-2
Нац. Метаб ., 2 , 127–129. DOI: 10.1038/s42255-020-0172-2 , Пякурель А., Глыцоу К. и Скоррано Л. (2020). Клеточная биология динамики митохондриальной мембраны. Нац. Преподобный Мол. Cell Biol ., 21 , 204–224. DOI: 10.1038/s41580-020-0210-7
, Пякурель А., Глыцоу К. и Скоррано Л. (2020). Клеточная биология динамики митохондриальной мембраны. Нац. Преподобный Мол. Cell Biol ., 21 , 204–224. DOI: 10.1038/s41580-020-0210-7 Comm ., 10 , 1287. DOI: 10.1038/s41467-019-09253-3
Comm ., 10 , 1287. DOI: 10.1038/s41467-019-09253-3