Оценка тремора при болезни Паркинсона с позиции теории мышечных веретен
Со времен К.Н. Третьякова [31] считается, что в основе болезни Паркинсона лежит гибель дофаминпродуцирующих нейронов черной субстанции среднего мозга [7, 12, 13, 20—22, 26, 28]. Дефицит церебрального дофамина приводит к тому, что хвостатое ядро не получает дофамин по нигростриарному пути и не срабатывает тормозное премоторно-каудато-паллидо-таламомоторное кольцо. Избыточная, т.е. ненужная в данный момент для деятельности мозга импульсация, приходящая от рецепторных полей организма, не тормозится и становится патологической [6, 7, 20, 21]. Она приходит в кору мозга, а затем по нисходящим экстрапирамидным путям, в том числе по рубро-ретикулоспинальному и нигро-ретикулоспинальному, достигает g-мотонейронов и реализует, в частности, дрожательный гиперкинез. Детали этой общей схемы будут уточнены после представления литературы и клинических и нейрофизиологических данных.
Известно, что в каждой крупной мышце выделяют волокна трех типов: I, IIА, IIБ [5, 14].
Медленные ядерносумочные интрафузальные мышечные волокна управляются преимущественно динамическими γ-мотонейронами, контролирующими динамическую стадию мышечного сокращения, при которой происходит изменение длины мышцы. Динамические γ-мотонейроны иннервируются, вероятно, нигроретикулоспинальным путем. Быстрые ядерноцепочечные интрафузальные мышечные волокна управляются преимущественно статическими γ-мотонейронами, контролирующими статическую фазу мышечного сокращения, при которой длина мышцы остается постоянной, например при изометрическом сокращении.
Влияние γ-мотонейронов и экстрапирамидной системы в целом на -мотонейроны осуществляется по моносинаптическому пути 1а от мышечных веретен к тоническим или фазическим α-мотонейронам. γ-Мотонейроны, мышечные веретена, путь 1а и α-мотонейроны образуют g-петлю, которая является анатомической основой миотатического рефлекса, или серво-механизма [4, 10, 11, 17], который играет ключевую роль в формировании мышечного тонуса и управлении произвольными и непроизвольными, в том числе насильственными движениями.
Цель работы — оценить дрожательно-ригидную форму болезни Паркинсона с позиций теории мышечных веретен.
Материал и методы
153 больным с дрожательно-ригидной формой болезни Паркинсона при относительно благоприятном течении сделано 197 стереотаксических операций. 3-я и 4-я стадии заболевания по Э.И. Канделю [7] при давности 6 лет и более, а также 1-я и 2-я стадии независимо от давности болезни дали основание диагностировать относительно благоприятное течение заболевания с медленным прогрессированием.
Стереотаксическими мишенями служили вентролатеральный ядерный комплекс таламуса, поля Н1, Н2 Фореля, неопределенная зона. Полное и почти полное прекращение тремора было достигнуто у 137 больных. Лучшие результаты (95% полного устранения или значительного уменьшения тремора) получены в группе мужчин с относительно благоприятным течением болезни Паркинсона.
Электромиографическое (ЭМГ) исследование (электромиограф Медикор) с помощью накожных электродов выполнено у 33 пациентов до операции и 14 — после. В качестве контроля для сравнения ЭМГ-показателей в динамике использовались ЭМГ-параметры мышц на «здоровой» стороне при гемипаркинсонизме. Исследованы дистальные (поверхностный сгибатель пальцев) и проксимальные (двуглавая мышца плеча) сгибатели руки, в качестве мышцы-разгибателя обследован общий разгибатель пальцев. ЭМГ записывали с режиме покоя и при функциональных пробах (вдох, синергия, сокращение и растяжение исследуемой мышцы, сокращение антагониста).
Результаты и обсуждение
Амплитуда ЭМГ отражает количественную сторону непроизвольного возбуждения фазических α-мотонейронов [8, 9, 16]. Рис. 1
Рис. 1
| Рис. 1. Амплитуда ЭМГ (в мкВ) сгибателя пальцев в покое и при функциональных пробах. |
| Таблица1 |
| ]]> |
 При пассивном растяжении двуглавой мышцы (сгибателя) плеча резко увеличена (р<0,01) амплитуда ЭМГ бицепса — до 295±73 мкВ. Пассивное растяжение бицепса ближе всего к прямой стимуляции веретен. Коэффициент реципрокности сгибателей, вычисленный по методике И.Г. Охнянской и А.А. Комаровой [18], составил 121±27% и показал нарушение реципрокных отношений между сгибателями и разгибателями.
При пассивном растяжении двуглавой мышцы (сгибателя) плеча резко увеличена (р<0,01) амплитуда ЭМГ бицепса — до 295±73 мкВ. Пассивное растяжение бицепса ближе всего к прямой стимуляции веретен. Коэффициент реципрокности сгибателей, вычисленный по методике И.Г. Охнянской и А.А. Комаровой [18], составил 121±27% и показал нарушение реципрокных отношений между сгибателями и разгибателями.Амплитуда ЭМГ разгибателя пальцев в покое составила 121±31 мкВ, превышая аналогичные показатели контроля и мышц-сгибателей. Глубокий вдох и синергия сопровождались увеличением высоты осцилляций до 180±40 мкВ. Коэффициент реципрокности составил 75±21%, указывая на то, что со стороны разгибателя отсутствуют грубые нарушения координационных отношений со сгибателями.
В целом оценка амплитуд ЭМГ указывала на высокой уровень непроизвольного возбуждения фазических α-мотонейронов сгибателей (см. рис. 1) и разгибателей, а также чувствительность α-мотонейронов к функциональным пробам, особенно проприоцептивную стимуляцию и пассивное растяжение бицепса.
ЭМГ-отражением тремора является залповая активность [15, 19, 23, 27, 29]. Залповый разряд, отведенный от поверхностного сгибателя пальцев в покое, продолжался 62±7 мс и отражал время непроизвольного возбуждения фазических α-мотонейронов (табл. 2).
| Таблица2 |
| ]]> |
 Пауза между залпами длилась 125±7 мс и соответствовала периоду торможения фазических α-мотонейронов или невозможности их возбуждения по закону «все или ничего». После фазы возбуждения нейрон погружается в состояние гиперполяризации и не отвечает на сигналы, приходящие от веретен по пути 1а в течение примерно 80 мс [17]. Согласно предположению H. Hufschmidt [30], при болезни Паркинсона имеет место продленная фаза гиперполяризации, которая определяет длительность паузы между залповыми разрядами в 125±7 мс.
Пауза между залпами длилась 125±7 мс и соответствовала периоду торможения фазических α-мотонейронов или невозможности их возбуждения по закону «все или ничего». После фазы возбуждения нейрон погружается в состояние гиперполяризации и не отвечает на сигналы, приходящие от веретен по пути 1а в течение примерно 80 мс [17]. Согласно предположению H. Hufschmidt [30], при болезни Паркинсона имеет место продленная фаза гиперполяризации, которая определяет длительность паузы между залповыми разрядами в 125±7 мс.| Рис. 2. Амплитуда ЭМГ (в мкВ) сгибателя во время пауз между залпами пальцев — в покое и при функциональных пробах. |
| ]]> |
 При дыхательной и синергической пробах был отмечена тенденция к увеличению амплитуды ЭМГ за редкими исключениями, но прирост амплитуды был менее заметен, чем в контрольных исследованиях. Это значит, что на интеро- и проприоцептивную стимуляцию реагировали нейроны с меньшей возбудимостью, чем в контрольных исследованиях. Во время произвольного сокращения и сокращения антагониста наблюдалось достоверное (р<0,01) возрастание амплитуды ЭМГ до 30,8±6,54—46,41±16,13 мкВ в сгибателях и 42,92±7,63—48,27±5,72 мкВ в разгибателе. Пассивное растяжение бицепса не изменило амплитуду ЭМГ по сравнению с состоянием покоя. Произвольное мышечное сокращение при болезни Паркинсона отличается тем, что на фазические α-мотонейроны одновременно воздействуют непроизвольное возбуждение от веретена по пути 1а и произвольная стимуляция, приходящая по пирамидным аксонам. В момент залпового разряда происходит суммация непроизвольного и произвольного возбуждения фазических α-мотонейронов. Во время паузы между залпами фазические α-мотонейроны, участвовавшие в формировании разряда, находятся в состоянии гиперполяризации и не отвечают как на произвольные, так и непроизвольные стимулы.
При дыхательной и синергической пробах был отмечена тенденция к увеличению амплитуды ЭМГ за редкими исключениями, но прирост амплитуды был менее заметен, чем в контрольных исследованиях. Это значит, что на интеро- и проприоцептивную стимуляцию реагировали нейроны с меньшей возбудимостью, чем в контрольных исследованиях. Во время произвольного сокращения и сокращения антагониста наблюдалось достоверное (р<0,01) возрастание амплитуды ЭМГ до 30,8±6,54—46,41±16,13 мкВ в сгибателях и 42,92±7,63—48,27±5,72 мкВ в разгибателе. Пассивное растяжение бицепса не изменило амплитуду ЭМГ по сравнению с состоянием покоя. Произвольное мышечное сокращение при болезни Паркинсона отличается тем, что на фазические α-мотонейроны одновременно воздействуют непроизвольное возбуждение от веретена по пути 1а и произвольная стимуляция, приходящая по пирамидным аксонам. В момент залпового разряда происходит суммация непроизвольного и произвольного возбуждения фазических α-мотонейронов. Во время паузы между залпами фазические α-мотонейроны, участвовавшие в формировании разряда, находятся в состоянии гиперполяризации и не отвечают как на произвольные, так и непроизвольные стимулы.
Нельзя оставить без внимания то, что статические γ-мотонейроны после избыточного возбуждения должны пройти фазу гиперполяризации, во время которой они не реагируют на избыточную импульсацию, приходящую по ретикуло-руброспинальным путям. В таком случае гиперполяризация γ-мотонейронов становится решающим фактором для прекращения залпового разряда на ЭМГ, а также возникновения и поддержания паузы между залповыми разрядами. Продленная гиперполяризация может быть объяснена частичным сложением периодов гиперполяризации статических γ-мотонейронов и фазических α-мотонейронов. Речь идет о частичном сложении указанных периодов: так как период гиперполяризации нейрона продолжается 80 мс [17], то удвоенный период гиперполяризации α- и γ-мотонейронов составил бы 160 мс, но в действительности пауза между разрядами продолжается 125±7 мс. При этом фаза гиперполяризации не совпадает по времени для статических γ-мотонейронов и фазических α-мотонейронов, а именно γ-мотонейроны вступают в фазу гиперполяризации и выходят из нее раньше, чем α-мотонейроны. После окончания фазы гиперполяризации для статического γ-мотонейрона он способен реагировать на избыточную импульсацию, приходящую по ретикуло-руброспинальному пути, что приводит к сокращению ядерноцепочечного интрафузального волокна и восстанавливает несоответствие в длине между интра- и экстрафузальной мышцами, так как интрафузальное волокно уже сокращено, а экстрафузальное — еще расслаблено под влиянием гиперполяризации фазических α-мотонейронов.
Продленная гиперполяризация может быть объяснена частичным сложением периодов гиперполяризации статических γ-мотонейронов и фазических α-мотонейронов. Речь идет о частичном сложении указанных периодов: так как период гиперполяризации нейрона продолжается 80 мс [17], то удвоенный период гиперполяризации α- и γ-мотонейронов составил бы 160 мс, но в действительности пауза между разрядами продолжается 125±7 мс. При этом фаза гиперполяризации не совпадает по времени для статических γ-мотонейронов и фазических α-мотонейронов, а именно γ-мотонейроны вступают в фазу гиперполяризации и выходят из нее раньше, чем α-мотонейроны. После окончания фазы гиперполяризации для статического γ-мотонейрона он способен реагировать на избыточную импульсацию, приходящую по ретикуло-руброспинальному пути, что приводит к сокращению ядерноцепочечного интрафузального волокна и восстанавливает несоответствие в длине между интра- и экстрафузальной мышцами, так как интрафузальное волокно уже сокращено, а экстрафузальное — еще расслаблено под влиянием гиперполяризации фазических α-мотонейронов. При болезни Паркинсона должно происходить синхронное и почти одновременное избыточное возбуждение пула статических γ-мотонейронов, после чего эти же мотонейроны одновременно оказываются в фазе гиперполяризации.
При болезни Паркинсона должно происходить синхронное и почти одновременное избыточное возбуждение пула статических γ-мотонейронов, после чего эти же мотонейроны одновременно оказываются в фазе гиперполяризации.
Указанный режим возбуждения и торможения передается фазическим α-мотонейронам, в результате чего от фазического оксидативного мышечного волокна отводятся залповые разряды с паузами между ними. Возможно, это связано с одновременным выключением тормозной функции соответствующего пула нейронов хвостатого ядра и функциональными свойствами самих статических γ-мотонейронов. После окончания фазы гиперполяризации для фазического α-мотонейрона он восстанавливает состояние поляризации и способен отвечать на сигналы, которые приходят по пути 1а от веретен иннервируемой им мышцы. К этому моменту интрафузальное волокно уже сокращено, а экстрафузальное — еще расслаблено. Возникают условия для очередного залпового разряда.
Нельзя не учитывать другие тормозные спинальные рефлексы, в частности аутогенное торможение с сухожильных рецепторов Гольджи и возвратное — через соответствующую коллатераль. Указанные тормозные рефлексы способны ослабить избыточное возбуждение α-мотонейронов и непроизвольное сокращение мышцы, тогда как решающий вклад в прекращение залпового разряда принадлежит механизму гиперполяризации нейронов, который определяет также длительность паузы между залповыми разрядами. В поддержании этой паузы, возможно, играет роль торможение со стороны веретен антагониста, так как в момент паузы в мышце-агонисте антагонист продуцирует залповый разряд. Если принять во внимание, что пауза между залпами сгибателя 125±7 мс почти вдвое продолжительнее залпового разряда разгибателя 69±8 мс (см. табл. 2), станет ясно, что механизм реципрокного торможения не может обеспечить столь длительную паузу между залповыми разрядами сгибателя. Следовательно, решающая роль в возникновении и поддержании паузы между залповыми разрядами принадлежит механизму продленной гиперполяризации фазических α-мотонейронов. Кроме того, в серии наших наблюдений встречались единичные записи, когда от сгибателя пальцев отводилась залповая активность, а в разгибателе пальцев залповых разрядов не было.
Указанные тормозные рефлексы способны ослабить избыточное возбуждение α-мотонейронов и непроизвольное сокращение мышцы, тогда как решающий вклад в прекращение залпового разряда принадлежит механизму гиперполяризации нейронов, который определяет также длительность паузы между залповыми разрядами. В поддержании этой паузы, возможно, играет роль торможение со стороны веретен антагониста, так как в момент паузы в мышце-агонисте антагонист продуцирует залповый разряд. Если принять во внимание, что пауза между залпами сгибателя 125±7 мс почти вдвое продолжительнее залпового разряда разгибателя 69±8 мс (см. табл. 2), станет ясно, что механизм реципрокного торможения не может обеспечить столь длительную паузу между залповыми разрядами сгибателя. Следовательно, решающая роль в возникновении и поддержании паузы между залповыми разрядами принадлежит механизму продленной гиперполяризации фазических α-мотонейронов. Кроме того, в серии наших наблюдений встречались единичные записи, когда от сгибателя пальцев отводилась залповая активность, а в разгибателе пальцев залповых разрядов не было. Подобный феномен приводят также И.Г. Охнянская и А.А. Комарова [18], B. Drechsler [29]. В указанных случаях пауза между залпами в сгибателе возникала без участия реципрокного механизма и могла поддерживаться за счет продленной гиперполяризации α-мотонейронов.
Подобный феномен приводят также И.Г. Охнянская и А.А. Комарова [18], B. Drechsler [29]. В указанных случаях пауза между залпами в сгибателе возникала без участия реципрокного механизма и могла поддерживаться за счет продленной гиперполяризации α-мотонейронов.
Дыхательная и синергическая пробы сопровождались достоверным (р<0,01) укорочением паузы между залпами в сгибателе пальцев в ответ на интроцептивную стимуляцию, что косвенно отражало усиление избыточного возбуждения фазических α-мотонейронов. Сокращение сгибателя и его антагониста приводило к угнетению залповой активности в сгибателе пальцев, что выразилось уменьшением числа записей, содержащих залпы с 17 в покое до 13 (см. табл. 2), а также укорочением паузы между залпами до 80±9—88±8 мс (р<0,01). Произвольная стимуляция при сокращении сгибателя сопровождалась вовлечением дополнительного пула мотонейронов, которые не участвовали в формировании залпов, и чьи разряды частично заполняли паузы между залпами. Сокращение антагониста (разгибатель пальцев) приводило к реципрокному торможениию фазических α-мотонейронов сгибателя, что также сокращало паузы между залпами.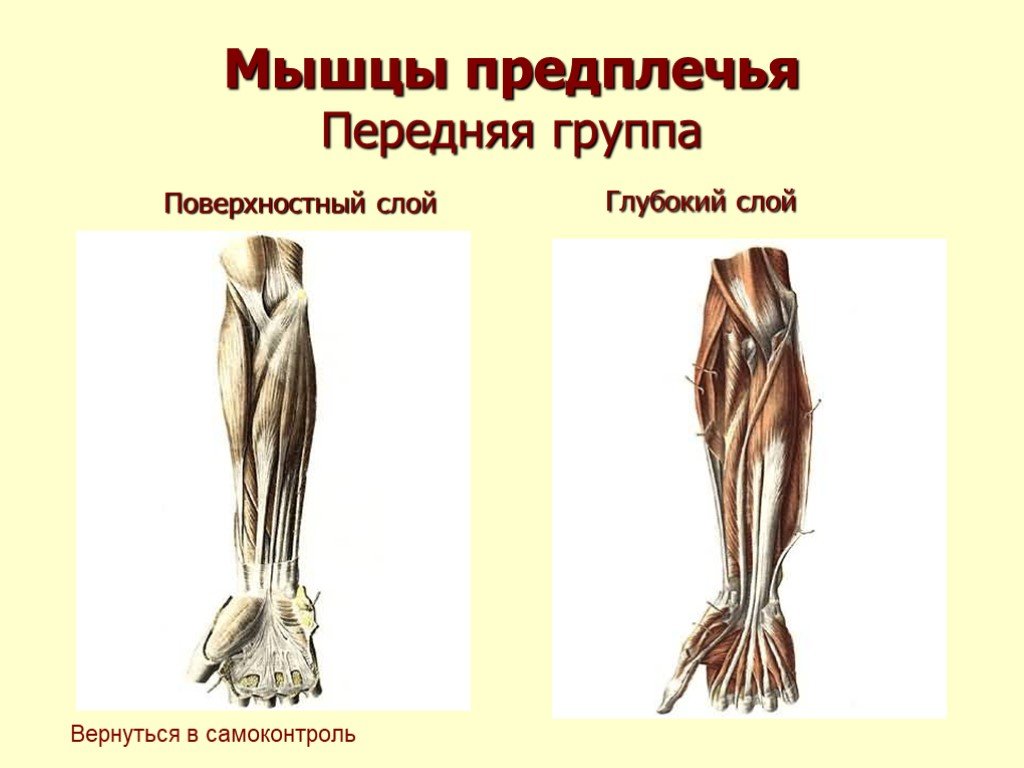 При этом ритм тремора участился до 7±0,3 в 1 с (р<0,01) по сравнению с состоянием покоя.
При этом ритм тремора участился до 7±0,3 в 1 с (р<0,01) по сравнению с состоянием покоя.
Любопытно сравнить характеристику проксимального и дистального сгибателя руки, т.е. двуглавой мышцы (сгибатель) плеча и поверхностного сгибателя пальцев (см. табл. 2). В покое в бицепсе отмечалось увеличение длительности одной осцилляции до 15±0,7 мс (р<0,05) по сравнению с аналогичным показателем сгибателя пальцев, что означает увеличение числа фазических мышечных волокон, вовлеченных в непроизвольное сокращение, и не противоречит представлению о том, что проксимальные мышцы человека имеют более крупные двигательные единицы по сравнению с дистальными. Длительность потенциала действия одной двигательной единицы бицепса при дрожательно-ригидной форме болезни Паркинсона составляет 7,3±0,53 мс [3], соответственно длительность одной осцилляции в структуре залпа может включать не менее двух двигательных единиц. Остальные показатели залповой активности, отведенной в состоянии покоя с проксимального и дистального сгибателей руки, не имели достоверных отличий.
Дыхательная и синергическая пробы по сравнению с состоянием покоя сопровождались увеличением (с 5 до 9) числа записей с залповой активностью, отведенной от бицепса, при этом количество осцилляций в залпе сокращалось с 5±0,8 до 3±0,4 (р<0,05) за счет уменьшения количества непроизвольно возбужденных фазических α-моторных единиц на фоне укорочения залпового разряда с 65±0,6 с до 53±9 — 57±0,5 мс. При глубоком вдохе длительность одной осцилляции также сокращалась с 15±0,7 до 13±1 мс (р<0,05), очевидно, за счет уменьшения числа мышечных волокон, подвергнутых непроизвольному сокращению. Длительность паузы между залпами не претерпела существенных изменений.
Произвольное сокращение бицепса сопровождалось увеличением количества записей с залповой активностью с 5 до 8 и увеличением длительности одной осцилляции с 15±0,7 до 16±0,01 мс (р<0,05), что означало увеличение числа сокращенных мышечных волокон, участвующих в залповом разряде, вероятно, за счет дополнительно включенных моторных единиц. Этому не противоречит тенденция к удлинению залпового разряда до 75±8 мс, однако число осцилляций в залпе имело тенденцию к уменьшению до 4±0,4. Пауза между залпами сократилась до 87±11 мс (р<0,02), чему не противоречило увеличение ритма тремора до 7±0,3 залпа в секунду.
Этому не противоречит тенденция к удлинению залпового разряда до 75±8 мс, однако число осцилляций в залпе имело тенденцию к уменьшению до 4±0,4. Пауза между залпами сократилась до 87±11 мс (р<0,02), чему не противоречило увеличение ритма тремора до 7±0,3 залпа в секунду.
По сравнению со сгибателем пальцев в разгибателе отмечалась тенденция к удлинению залпового разряда, укорочению паузы между залпами, уменьшению длительности одной осцилляции (см. табл. 2). Отмечено учащение ритма дрожания до 7±0,4 в 1 с (р<0,01). При вдохе по сравнению с состоянием покоя имело место удлинение одной осцилляции до 12±0,05 мс (р<0,05) за счет увеличения числа непроизвольно сокращенных фазических мышечных волокон на фоне тенденции к удлинению залпового разряда до 76±6 мс. Число осцилляций в залпе увеличилось до 6±0,6 (р<0,05) за счет увеличения непроизвольно возбужденных фазических двигательных единиц. Пауза между залпами имела тенденцию к увеличению до 106±9 мс. Ритм залповых разрядов снизился до 6±0,4 (р<0,05).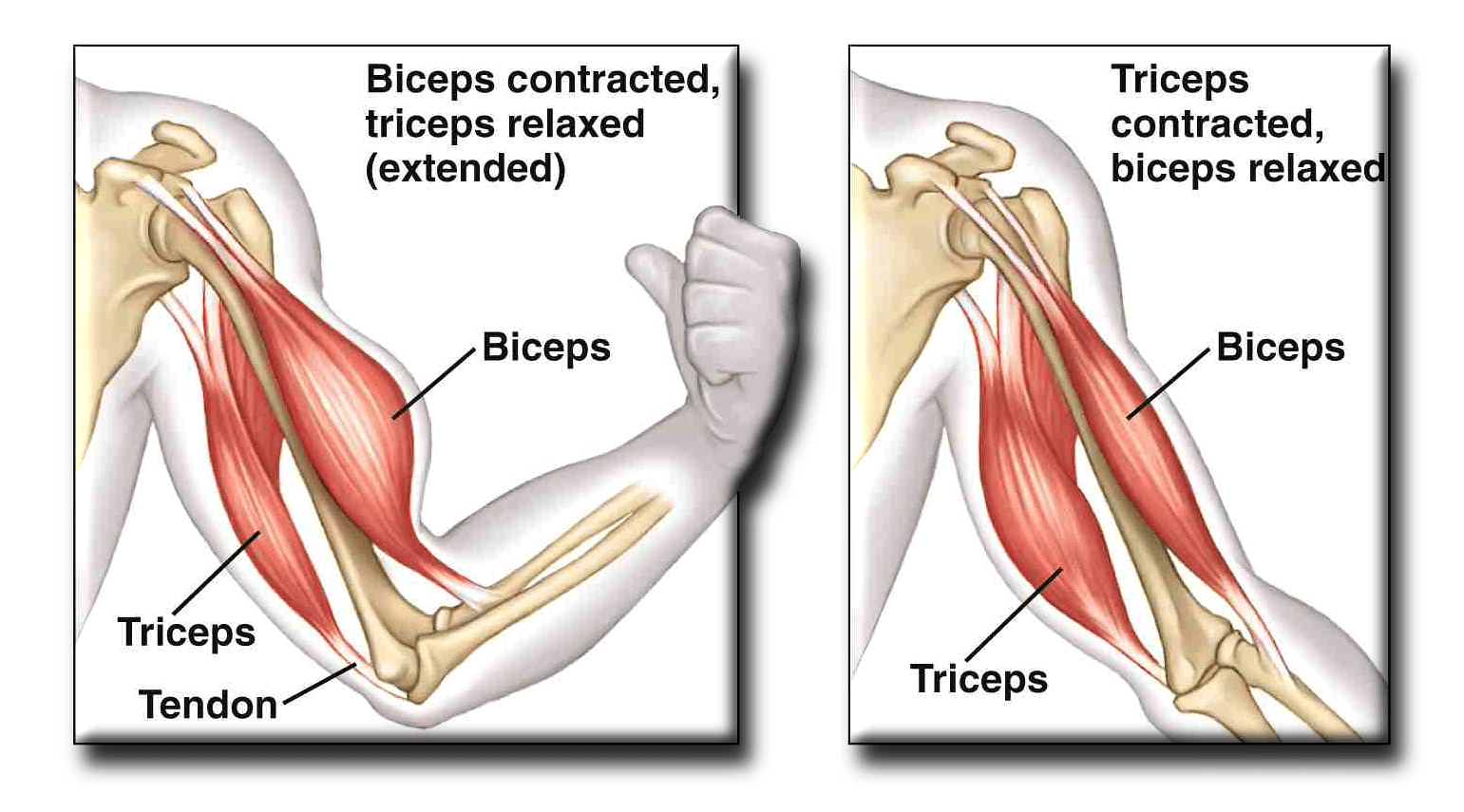
Следовательно, продолжительность залпового разряда в сгибателях короче, а пауза между залпами длиннее, чем в разгибателе. Приведенный факт свидетельствует о преобладании избыточного возбуждения в -мотонейронах разгибателей, вероятно, за счет антигравитационной нагрузки, что подтверждается также более высокой амплитудой ЭМГ разгибателя (см. табл. 1).
У единичных больных залповая активность была нерегулярной. Ее анализ представляет интерес, поскольку нерегулярная залповая активность предшествует формированию регулярных залпов и позволяет уточнить некоторые особенности формирования тремора. Наиболее часто она отводилась от мышц рук сгибательной группы. Возможно, сгибатели в большей мере, чем разгибатели, «противодействуют» формированию залповой активности. Чаще всего нерегулярную залповую активность провоцировали произвольное сокращение агониста и особенно — антагониста; наиболее редко нерегулярные залповые разряды отводили от мышц в состоянии физиологического покоя. По-видимому, в происхождении залповых разрядов играет роль импульсация от веретен, так как сокращение антагониста сопровождается растяжением агониста, при этом происходит раздражение сенсорных окончаний веретена.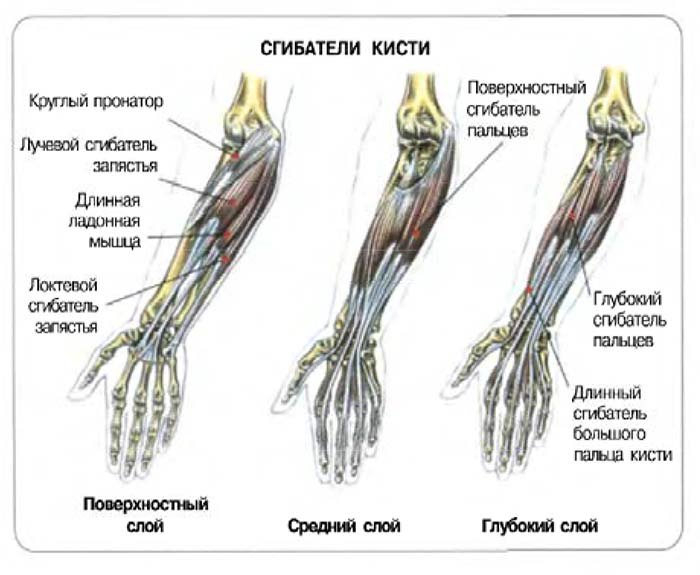 Указанному факту не противоречит возрастание числа записей с нерегулярной залповой активностью от бицепса при его пассивном растяжении до 4 против 1 записи в состоянии покоя.
Указанному факту не противоречит возрастание числа записей с нерегулярной залповой активностью от бицепса при его пассивном растяжении до 4 против 1 записи в состоянии покоя.
После операции залповая активность устранялась, что объективно подтверждало устранение дрожательного гиперкинеза. У единичных больных от некоторых мышц сгибательной группы в покое, а также при дыхательной пробе отводились нерегулярные залповые разряды. Длительность залпового разряда составила 25—75 мс (табл. 3).
| Таблица3 |
| ]]> |

Наиболее полно залповые разряды были оформлены в общем разгибателе пальцев. В покое, при вдохе и синергической пробе они укорачивались до 25 — 75 мс по сравнению с дооперационным уровнем, чему соответствовало уменьшение числа осцилляций в залпе до трех, что указывало на уменьшение количества двигательных единиц, участвующих в формировании залпа. Продолжительность одной осцилляции составила 8—16 мс, пауза между залпами — 100 мс. Ритм залповых разрядов после операции не изменился. Сокращение разгибателя пальцев и его антагониста привело к укорочению одной осцилляции до 8 мс, остальные показатели залповой активности существенно не изменились (см. табл. 3).
Следовательно, перестройка остаточной залповой активности после операции заключалась в укорочении залповых разрядов, уменьшении числа осцилляций в залпе, что свидетельствовало об уменьшении непроизвольно возбужденных двигательных единиц. Тенденция к укорочению одной осцилляции указывала на уменьшение количества непроизвольно сокращенных фазических мышечных волокон, участвующих в формировании тремора. Хуже всего залповая активность устранялась в разгибателе пальцев, очевидно, за счет более четкого реагирования разгибательных мышц на гравитационную нагрузку. Кроме того, после операции и сгибатели, и разгибатели охотнее продуцировали залповую активность в ответ на произвольное сокращение и сокращение антагониста, что могло указывать на преимущественную роль проприорецепторов, т.е. веретен в происхождении тремора. Обращало внимание некоторое сходство нерегулярной и остаточной залповой активности после операции.
Хуже всего залповая активность устранялась в разгибателе пальцев, очевидно, за счет более четкого реагирования разгибательных мышц на гравитационную нагрузку. Кроме того, после операции и сгибатели, и разгибатели охотнее продуцировали залповую активность в ответ на произвольное сокращение и сокращение антагониста, что могло указывать на преимущественную роль проприорецепторов, т.е. веретен в происхождении тремора. Обращало внимание некоторое сходство нерегулярной и остаточной залповой активности после операции.
После операции амплитуда ЭМГ сгибателей в покое (13±1,7—19±0,4 мкВ) была достоверно (р<0,01) меньше (см. табл. 1), что указывало на снижение избыточной возбудимости фазических α-мотонейронов. Дыхательная и синергическая пробы приводили к достоверному (р<0,01) снижению высоты осцилляций до 17±5,7—41±25 мкВ по сравнению с дооперационными показателями, однако они были выше, чем в состоянии функционального покоя. Следовательно, α-мотонейроны сохраняли способность реагировать на функциональные раздражители (см. рис. 1). Произвольное сокращение сгибателей и проба на реципрокность иннервации не вызывали достоверного изменения амплитуды ЭМГ, что указывало на отсутствие послеоперационных парезов.
рис. 1). Произвольное сокращение сгибателей и проба на реципрокность иннервации не вызывали достоверного изменения амплитуды ЭМГ, что указывало на отсутствие послеоперационных парезов.
После операции амплитуда ЭМГ разгибателей в состоянии покоя достоверно (р<0,02) уменьшилась до 13±3,7—38±17 мкВ. Глубокий вдох и синергия сопровождались снижением (р<0,01) высоты осцилляций разгибателей до 21±7—42±23 мкВ. По сравнению с состоянием покоя имелась тенденция к увеличению уровня возбудимости фазических α-мотонейронов. Произвольное сокращение самих разгибателей и сокращение их антагонистов провоцировали тенденцию к некоторому росту амплитуды ЭМГ разгибателей (см. табл. 1). Коэффициенты реципрокности и адекватности разгибателя имели тенденцию к уменьшению.
В целом динамика амплитуд мышечных потенциалов убедительно показывает снижение непроизвольного возбуждения фазических α-мотонейронов и объективно подтверждает лечебный эффект хирургического лечения.
Заключение
На основании представленных данных выстраивается гипотеза происхождения тремора, которая может быть использована для понимания других гиперкинезов [24, 25]. Энергетической базой гиперкинезов является естественная афферентная импульсация с рецептивных полей организма, особенно от проприорецепторов, или мышечных веретен. В стволе мозга часть этой импульсации переключается на ядра ретикулярной формации и по неспецифическим путям в составе восходящей активирующей системы достигает ассоциативной коры большого мозга, где используется для поддержания бодрствования и внимания [2]. Известно, что в состоянии сна восходящая активирующая система выключается, что сопровождается устранением тремора и других гиперкинезов.
Энергетической базой гиперкинезов является естественная афферентная импульсация с рецептивных полей организма, особенно от проприорецепторов, или мышечных веретен. В стволе мозга часть этой импульсации переключается на ядра ретикулярной формации и по неспецифическим путям в составе восходящей активирующей системы достигает ассоциативной коры большого мозга, где используется для поддержания бодрствования и внимания [2]. Известно, что в состоянии сна восходящая активирующая система выключается, что сопровождается устранением тремора и других гиперкинезов.
Бодрствующий мозг отводит избыточную неспецифическую импульсацию по тормозному премоторно-каудато-паллидо-таламомоторному пути (каудатная петля), которое функционирует по принципу отрицательной обратной связи. Устранение избыточных импульсов осуществляется на уровне хвостатого ядра с помощью дофамина, который поступает в хвостатое ядро по нигростриарному пути. При болезни Паркинсона каудатная петля не срабатывает из-за дефицита церебрального дофамина. При хорее Гентингтона, торсионной дистонии и атетозе имеет место первичное гнездное поражение нейронов хвостатого ядра («мраморное состояние»).
При хорее Гентингтона, торсионной дистонии и атетозе имеет место первичное гнездное поражение нейронов хвостатого ядра («мраморное состояние»).
Следует допустить, что в хвостатом ядре существуют популяции нейронов для избирательного автономного воздействия (управление) на веретена, принадлежащие либо тоническим типа I, либо оксидативным фазическим типа IIa, либо гликолитическим фазическим типа IIb мышечным волокнам. Это управление осуществляется по полисинаптическим экстрапирамидным путям, включая γ-мотонейроны. В результате поражения хвостатого ядра избыточная импульсация не гасится и становится гиперкинезогенной. На пути к моторной коре через каудатную петлю она проходит через вентролатеральный ядерный комплекс таламуса, что доказано устранением гиперкинеза в результате стереотаксических операций на зрительном бугре. Нетормозимая избыточная гиперкинезогенная импульсация по каудатной петле поступает в моторную кору, откуда по полисинаптическим экстрапирамидным путям, включая корково-мостомозжечково-зубчато-красноядерно-ретикулоспинальный и др. , достигает γ-мотонейронов [1].
, достигает γ-мотонейронов [1].
Считается, что при болезни Паркинсона избирательно страдает управление статическими γ-мотонейронами. Далее необходимо сделать допущение, что происходит одновременное синхронное избыточное возбуждение большого пула статических γ-мотонейронов. После возбуждения этот же пул впадает в состояние гиперполяризации и не отвечает на сигналы, приходящие по ретикуло-руброспинальному пути. Указанное синхронное избыточное возбуждение статических γ-мотонейронов и последующее их синхронное погружение в фазу гиперполяризации является ключевым моментом в части спинальных механизмов тремора. Избыточное возбуждение статических γ-мотонейронов приводит к сокращению ядерноцепочечных интрафузальных мышечных волокон, принадлежащих фазическим оксидативным экстрафузальным волокнам типа IIa, затем срабатывает серво-механизм, и по моносинаптическому пути 1а патологический сигнал без искажения передается фазическому α-мотонейрону. Последний сокращает экстрафузальное мышечное волокно типа IIА, и на ЭМГ возникает залповый разряд, характеристика которого обсуждена в тексте статьи.
Таким образом, на спинальном уровне дрожательный гиперкинез реализует миотатическая единица с участием статического γ-мотонейрона, ядерноцепочечного интрафузального волокна, пути 1а, фазических α-мотонейрона и оксидативного экстрафузального мышечного волокна типа IIА. Представленная гипотеза позволяет глубже понять патогенез тремора, а также оценить результаты хирургического лечения.
Упражнения на массу сгибателей и пронаторов выборочно активизируют мускулатуру предплечья
Выбросы из-за головы создают чрезмерную вальгусную нагрузку на медиальную часть локтя. 1,2 Это вальгусное напряжение преимущественно возникает во время поздних фаз взведения и ускорения метания и в первую очередь сопротивляется переднему пучку локтевой коллатеральной связки (ЛКС). 2,3 Эта повторяющаяся нагрузка на ВКС считается фактором, способствующим увеличению частоты разрывов ВКС. 4
Максимальная сила, которой может противостоять UCL, составляет около 54% от вальгусной силы, возникающей при качке. 2 Таким образом, постулируется, что динамическая стабильность локтевого сустава обеспечивается сгибательно-пронаторной мускулатурой медиальной части локтевого сустава. 5–11 В частности, локтевой сгибатель запястья (FCU), поверхностный сгибатель пальцев (FDS) и круглый пронатор (PT) были предложены в качестве активных стабилизаторов локтевого сустава во время вальгусной нагрузки. 5,6,8,10,11 Это предположение основано на анатомическом расположении массы сгибателей-пронаторов в непосредственной близости от UCL. 5,10 Также было показано, что изометрическое сокращение медиальной мускулатуры предплечья уменьшает пространство плече-локтевого сустава во время вальгусной нагрузки 11 , а медиальная стабильность локтевого сустава снижается при утомлении мускулатуры предплечья, 12 , что еще больше указывает на их динамику вклад в стабильность.
2 Таким образом, постулируется, что динамическая стабильность локтевого сустава обеспечивается сгибательно-пронаторной мускулатурой медиальной части локтевого сустава. 5–11 В частности, локтевой сгибатель запястья (FCU), поверхностный сгибатель пальцев (FDS) и круглый пронатор (PT) были предложены в качестве активных стабилизаторов локтевого сустава во время вальгусной нагрузки. 5,6,8,10,11 Это предположение основано на анатомическом расположении массы сгибателей-пронаторов в непосредственной близости от UCL. 5,10 Также было показано, что изометрическое сокращение медиальной мускулатуры предплечья уменьшает пространство плече-локтевого сустава во время вальгусной нагрузки 11 , а медиальная стабильность локтевого сустава снижается при утомлении мускулатуры предплечья, 12 , что еще больше указывает на их динамику вклад в стабильность.
Медиальная мускулатура предплечья также демонстрирует высокую электромиографическую (ЭМГ) активность во время броскового движения, особенно во время поздней фазы взведения и раннего ускорения броска. 13–16 Ранее было продемонстрировано, что в этих фазах качка возникает максимальное вальгусное напряжение медиальной части локтя, 2 , что побудило исследователей предположить, что сгибательно-пронаторная масса обеспечивает динамическую стабилизацию медиальной части локтя. 13–15 Кроме того, мускулатура предплечья считается ключевым компонентом точности подачи и движения мяча, что еще раз подтверждает важность мышц-сгибателей-пронаторов для спортсмена, выполняющего метание. 17,18
13–16 Ранее было продемонстрировано, что в этих фазах качка возникает максимальное вальгусное напряжение медиальной части локтя, 2 , что побудило исследователей предположить, что сгибательно-пронаторная масса обеспечивает динамическую стабилизацию медиальной части локтя. 13–15 Кроме того, мускулатура предплечья считается ключевым компонентом точности подачи и движения мяча, что еще раз подтверждает важность мышц-сгибателей-пронаторов для спортсмена, выполняющего метание. 17,18
Основываясь на роли мышц-сгибателей-пронаторов во время броскового движения, укрепление этих мышц считается важным компонентом программы реабилитации для спортсменов, занимающихся прыжками в высоту, и потенциально полезно для смягчения травм UCL. 5,6,8,16,19 На сегодняшний день недостаточно данных, чтобы предложить конкретные вмешательства, направленные на сгибательно-пронаторную мускулатуру предплечья. Сопротивление эластичной ленты представляет собой портативное и удобное средство для укрепления мышц-сгибателей-пронаторов. Однако о степени активности ЭМГ мускулатуры сгибателей-пронаторов при использовании эластического сопротивления не сообщалось. Таким образом, целью данного исследования было измерение степени активности ЭМГ мышц-сгибателей-пронаторов во время двух различных упражнений на укрепление предплечья с использованием эспандеров. Было высказано предположение, что два упражнения вызовут, по крайней мере, умеренный уровень мышечной активности, но активация пронаторов и мышц-сгибателей будет отличаться 9.0009
Однако о степени активности ЭМГ мускулатуры сгибателей-пронаторов при использовании эластического сопротивления не сообщалось. Таким образом, целью данного исследования было измерение степени активности ЭМГ мышц-сгибателей-пронаторов во время двух различных упражнений на укрепление предплечья с использованием эспандеров. Было высказано предположение, что два упражнения вызовут, по крайней мере, умеренный уровень мышечной активности, но активация пронаторов и мышц-сгибателей будет отличаться 9.0009
Участники
Десять субъектов мужского пола были набраны в качестве удобной выборки из местного сообщества. Все испытуемые были здоровы на момент тестирования, не имели в анамнезе патологии локтя или запястья, и было подтверждено, что у них нормальный функциональный диапазон движений в доминирующем запястье и предплечье, и они были в состоянии выполнять задачи, включенные в это исследование. . Восемь из десяти испытуемых участвовали в регулярных силовых тренировках не менее трех раз в неделю; однако ни один из испытуемых не сообщил о том, что выполнял укрепляющие упражнения, направленные конкретно на мускулатуру предплечья. Четверо из десяти испытуемых в анамнезе занимались метанием или спортом над головой. Перед участием в этом исследовании каждый субъект предоставил письменное информированное согласие в соответствии с правилами институционального наблюдательного совета (Northwell Health IRB # 19).-0586)
Четверо из десяти испытуемых в анамнезе занимались метанием или спортом над головой. Перед участием в этом исследовании каждый субъект предоставил письменное информированное согласие в соответствии с правилами институционального наблюдательного совета (Northwell Health IRB # 19).-0586)
Таблица 1. Характеристика субъекта.
| Количество субъектов | 10 (все мужские) |
| Возраст | 36 ± 12 лет |
| Высота | 1,77 ± 0,04 м |
| Вес | 79 ± 5 кг |
Среднее значение ± стандартное отклонение
Процедуры
16-канальная система BTS FREEEMG 300, КОСС: >110 дБ при 50–60 Гц; входное сопротивление: >10 ГОм (BTS Bioengineering, Милан, Италия) использовалось для сбора ЭМГ во время этого исследования.
Ранее описанные анатомические ориентиры для размещения поверхностной ЭМГ были определены для мышц FCU, FDS и PT. 20,21 Открытая кожа субъекта была подготовлена путем бритья, очистки и легкого натирания. Одноразовые пассивные ЭМГ-электроды с двойной поверхностью Ag/AgCl (расстояние между электродами 2,0 см; Noraxon, Скоттсдейл, Аризона) были помещены на определенные ориентиры на правом проксимальном предплечье каждого субъекта (рис. 1). Мышечная активность измерялась при частоте 1000 Гц.
20,21 Открытая кожа субъекта была подготовлена путем бритья, очистки и легкого натирания. Одноразовые пассивные ЭМГ-электроды с двойной поверхностью Ag/AgCl (расстояние между электродами 2,0 см; Noraxon, Скоттсдейл, Аризона) были помещены на определенные ориентиры на правом проксимальном предплечье каждого субъекта (рис. 1). Мышечная активность измерялась при частоте 1000 Гц.
Рисунок 1. Расположение электродов ЭМГ: (A) круглый пронатор, (B) поверхностный сгибатель пальцев, (C) 9-й локтевой сгибатель запястья0009
После прикрепления поверхностных электродов к субъекту выполнялось максимальное произвольное сокращение (MVC) в ранее описанном положении мануального мышечного тестирования (MMT) для каждой мышцы. 22 Один испытатель, физиотерапевт с более чем 20-летним клиническим опытом, провел ММТ для всех испытуемых.
Затем испытуемые определяли уровень сопротивления, который они будут использовать в своем испытании, выполняя пятикратное отклонение запястья и локтевой кости, удерживая эластичную ленту (без латекса TheraBand, Hygenic Corporation, Акрон, Огайо). Лента сопротивления была обрезана до длины ноги каждого субъекта, чтобы обеспечить дополнительную длину для обертывания вокруг руки. Чтобы стандартизировать уровень сопротивления для двух упражнений, испытуемые определяли, какой уровень сопротивления вызывал нагрузку 5 из 10 по шкале Борга CR10. 23 Этот процесс был повторен для пронации предплечья. Каждому испытуемому были даны инструкции, чтобы в начале упражнения не было натяжения или провисания эспандера, поскольку удлинение эспандера влияет на воспринимаемое сопротивление и связанную с ним мышечную активность. 24
Лента сопротивления была обрезана до длины ноги каждого субъекта, чтобы обеспечить дополнительную длину для обертывания вокруг руки. Чтобы стандартизировать уровень сопротивления для двух упражнений, испытуемые определяли, какой уровень сопротивления вызывал нагрузку 5 из 10 по шкале Борга CR10. 23 Этот процесс был повторен для пронации предплечья. Каждому испытуемому были даны инструкции, чтобы в начале упражнения не было натяжения или провисания эспандера, поскольку удлинение эспандера влияет на воспринимаемое сопротивление и связанную с ним мышечную активность. 24
Два упражнения: локтевая девиация запястья и пронация предплечья выполнялись в случайном порядке. Эти упражнения были включены в программу по уходу за руками, основанную на доказательствах, ранее опубликованную со ссылками на видеоролики с упражнениями. 25 Рандомизация проводилась путем извлечения карточки из непрозрачного конверта с указанием упражнения, написанного на выбранной карточке, которое нужно выполнить первым.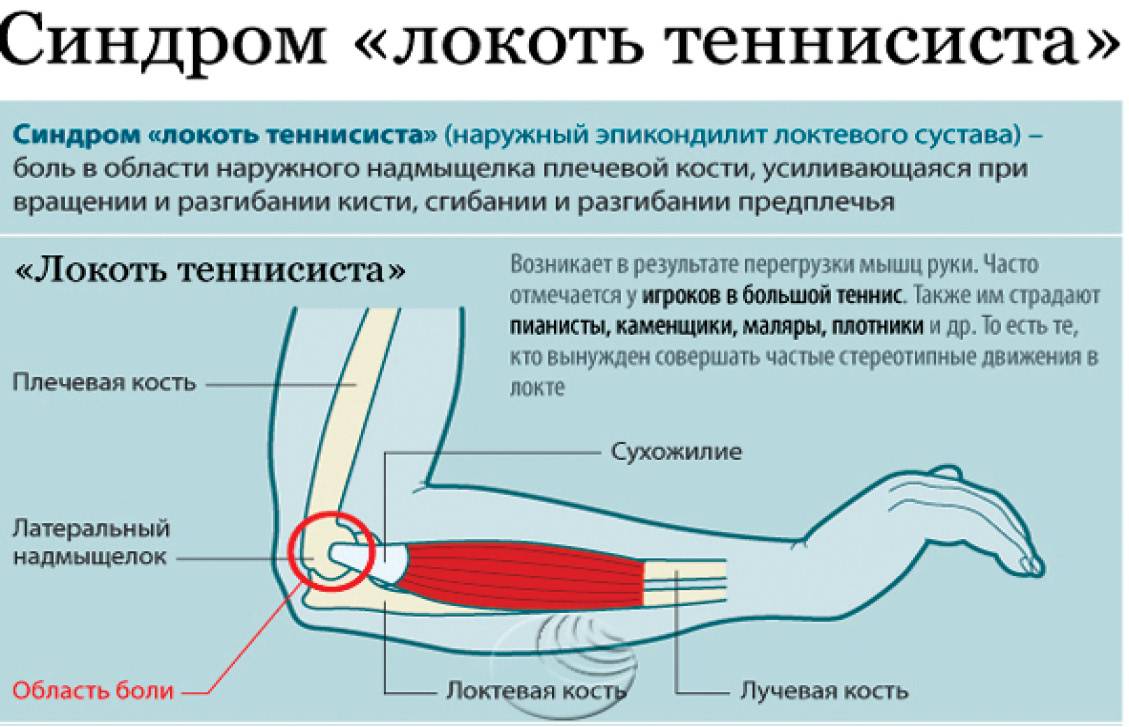 Ульнарное отклонение запястья выполнялось испытуемым, держащим эспандер с вытянутыми локтями, ладонями вниз и руками на уровне плеч. Эластичная лента была дважды обернута вокруг рук испытуемого, при этом лента не провисала и не натягивалась, когда руки находились на ширине плеч (рис. 2А). Затем испытуемый переместил запястье в локтевую девиацию (рис. 2В), выполнив это движение три раза. Субъекта просили сопоставить скорость движения с метрономом, установленным на 20 ударов в минуту, направляя субъекта на выполнение 3-секундной концентрической фазы и 3-секундной эксцентрической фазы, чтобы обеспечить контролируемое движение в обеих фазах упражнения.
Ульнарное отклонение запястья выполнялось испытуемым, держащим эспандер с вытянутыми локтями, ладонями вниз и руками на уровне плеч. Эластичная лента была дважды обернута вокруг рук испытуемого, при этом лента не провисала и не натягивалась, когда руки находились на ширине плеч (рис. 2А). Затем испытуемый переместил запястье в локтевую девиацию (рис. 2В), выполнив это движение три раза. Субъекта просили сопоставить скорость движения с метрономом, установленным на 20 ударов в минуту, направляя субъекта на выполнение 3-секундной концентрической фазы и 3-секундной эксцентрической фазы, чтобы обеспечить контролируемое движение в обеих фазах упражнения.
Рисунок 2. Локтевое отклонение запястья (A) исходное положение (B) конечное положение.
Пронация предплечья выполнялась в положении обследуемого сидя. Лента сопротивления была дважды обернута вокруг руки. Затем испытуемый натягивал эспандер до середины голени и наступал на эспандер правой ногой. Их предплечье было помещено на правое бедро в супинированном положении, правая складка запястья находилась на краю надколенника. (Рисунок 3А). Испытуемому было дано указание выполнить весь доступный диапазон пронационных движений предплечья в трех повторениях (рис. 3В). Скорость движения соответствовала метроному, установленному на 20 ударов в минуту, что приводило к 3-секундной концентрической фазе и 3-секундной эксцентрической фазе.
(Рисунок 3А). Испытуемому было дано указание выполнить весь доступный диапазон пронационных движений предплечья в трех повторениях (рис. 3В). Скорость движения соответствовала метроному, установленному на 20 ударов в минуту, что приводило к 3-секундной концентрической фазе и 3-секундной эксцентрической фазе.
Рисунок 3. Пронация предплечья (A) исходное положение (B) конечное положение.
Обработка данных
Все данные ЭМГ были обработаны в соответствии с ранее описанными методами. 26 Вкратце, необработанные сигналы ЭМГ прошли фильтрацию верхних частот на частоте 10 Гц, двухполупериодное выпрямление и сглаживание с использованием среднеквадратичного фильтра с окном 100 мс. Для каждой мышцы определяли пиковый уровень активации ЭМГ во время каждого упражнения. Затем рассчитывали средний уровень активации каждой мышцы от 250 миллисекунд до пика до 250 миллисекунд после пика. Затем это значение усреднялось по трем повторениям для каждого упражнения и каждой мышцы. После обработки данные ЭМГ были нормализованы к максимальной активности ЭМГ, зарегистрированной во время максимальных произвольных сокращений (МПС), и выражены в процентах. Уровень мышечной активности классифицировали по системе DiGiovin et al. как от 0% до 20% низкой активности MVC, от 21% до 40% умеренной активности MVC, от 41% до 60% высокой активности MVC и более 60% очень высокой активности MVC. 14
После обработки данные ЭМГ были нормализованы к максимальной активности ЭМГ, зарегистрированной во время максимальных произвольных сокращений (МПС), и выражены в процентах. Уровень мышечной активности классифицировали по системе DiGiovin et al. как от 0% до 20% низкой активности MVC, от 21% до 40% умеренной активности MVC, от 41% до 60% высокой активности MVC и более 60% очень высокой активности MVC. 14
Описательная статистика была рассчитана для средней нормализованной пиковой мышечной активности для каждой мышцы во время каждого упражнения. Влияние упражнений и мышц на среднюю нормализованную пиковую активность ЭМГ сравнивали с использованием двухфакторного (упражнение x мышца) повторных измерений ANOVA. Если эффект взаимодействия был обнаружен, рассчитывались апостериорные простые главные эффекты с поправкой Бонферрони. Уровень значимости был установлен априори при р = 0,05. Все статистические анализы были выполнены с использованием IBM SPSS Statistics 25 (IBM Corp, Армонк, Нью-Йорк).
Средняя нормализованная мышечная активность для каждой мышцы во время каждого изучаемого упражнения показана в таблице 2. Было проведено упражнение по эффекту взаимодействия мышц (p < 0,001). Упражнение на отклонение локтевой кости избирательно активировало FCU (нормализованная пиковая ЭМГ 40,3%) по сравнению с FDS (19,5%, p = 0,009) и PT (21,5%, p = 0,022). Согласно системе классификации DiGiovine et al., упражнения с отклонением локтевой кости вызывали умеренную мышечную активность при FCU, низкую активность при FDS и умеренную активность при PT. И наоборот, упражнение на пронацию избирательно активировало ФДС (63,9%, p = 0,002) и PT (73,0 %, p = 0,001) по сравнению с FCU (27,4 %). Упражнение с пронацией вызывало очень высокую мышечную активность при FDS и PT и умеренную мышечную активность при FCU.
Таблица 2. Средняя мышечная активность для исследуемых мышц и упражнений.
| Упражнение на отклонение локтевой кости | Упражнение на пронацию | |
| Локтевой сгибатель запястья (FCU) | 40,3 ± 20,2* | 27,4 ± 11,7** |
| Поверхностный сгибатель пальцев (FDS) | 19,5 ± 18,0* | 63,9 ± 31,4** |
| Круглый пронатор (PT) | 21,5 ± 19,9* | 73,0 ± 32,1** |
Все значения являются средними % MVC ± SD
* FCU значительно выше активность, чем FDS (p = 0,009) и PT (p = 0,022)
** FDS (p = 0,002) и PT (p = 0,001) значительно выше активность, чем у FCU
. Результаты текущего исследования показывают, что упражнения с использованием сопротивления эластичных лент избирательно нацелены и вызывают активность от умеренной до очень высокой в мышцах-сгибателях-пронаторах. Нормализованная пиковая ЭМГ была значительно выше в FCU по сравнению с FDS и PT во время упражнения на отклонение локтевой кости, в то время как нормализованная пиковая EMG была значительно выше в FDS и PT во время упражнения на пронацию. Следовательно, FCU преимущественно активировался во время упражнения на отклонение локтевой кости, в то время как FDS и PT были более активны во время упражнения на пронацию.
Результаты текущего исследования показывают, что упражнения с использованием сопротивления эластичных лент избирательно нацелены и вызывают активность от умеренной до очень высокой в мышцах-сгибателях-пронаторах. Нормализованная пиковая ЭМГ была значительно выше в FCU по сравнению с FDS и PT во время упражнения на отклонение локтевой кости, в то время как нормализованная пиковая EMG была значительно выше в FDS и PT во время упражнения на пронацию. Следовательно, FCU преимущественно активировался во время упражнения на отклонение локтевой кости, в то время как FDS и PT были более активны во время упражнения на пронацию.
Масса мышц-сгибателей и пронаторов является важной группой мышц у спортсменов, занимающихся верхом, так как она обеспечивает динамическую стабильность локтя для противодействия вальгусным силам. 5–11 Эта динамическая стабилизация имеет решающее значение, так как качка создает большую нагрузку на ВКЛ, 1,2 и скорость разрыва ВКЛ возрастает. 4 Кроме того, мышцы-сгибатели-пронаторы имеют высокий уровень активности во время бросков из-за головы 13–16 и важны для контроля и скорости подачи. 17,18 Несмотря на их роль в бросках из-за головы, существует минимальное количество данных о том, как эффективно тренировать эти мышцы и можно ли направить на них укрепляющие упражнения.
4 Кроме того, мышцы-сгибатели-пронаторы имеют высокий уровень активности во время бросков из-за головы 13–16 и важны для контроля и скорости подачи. 17,18 Несмотря на их роль в бросках из-за головы, существует минимальное количество данных о том, как эффективно тренировать эти мышцы и можно ли направить на них укрепляющие упражнения.
Информация о том, что эти упражнения избирательно активируют мышцы-сгибатели-пронаторы, важна для спортсменов, работающих над головой, а также для силовых, кондиционно-реабилитационных специалистов из-за возможности повышения динамической стабильности локтевого сустава и потенциального снижения нагрузки на UCL. 5,6,8,16,19 Эти результаты могут повлиять на принятие клинического решения, поскольку для укрепления всех мышц сгибателей-пронаторов и наилучшего укрепления динамических стабилизаторов медиальной части локтевого сустава требуется более одного упражнения.
У этого исследования было несколько ограничений. В выборку исследования входили только здоровые участники мужского пола, и это ограничивает возможность обобщения его результатов, особенно для пациентов с травмами, поскольку у пациентов с травмой UCL в анамнезе наблюдаются изменения мышечной активности. 19 Поперечный характер этого исследования не позволяет изучить тренировочный эффект этих упражнений. Необходимы дальнейшие исследования для выяснения их тренировочного воздействия на мышечную силу, активацию мышц во время бросков из-за головы и медиальную стабильность локтя.
В выборку исследования входили только здоровые участники мужского пола, и это ограничивает возможность обобщения его результатов, особенно для пациентов с травмами, поскольку у пациентов с травмой UCL в анамнезе наблюдаются изменения мышечной активности. 19 Поперечный характер этого исследования не позволяет изучить тренировочный эффект этих упражнений. Необходимы дальнейшие исследования для выяснения их тренировочного воздействия на мышечную силу, активацию мышц во время бросков из-за головы и медиальную стабильность локтя.
Результаты текущего исследования показывают, что упражнения на отклонение локтевой кости и пронацию с использованием эластичных лент могут нацеливаться и активировать массу сгибателей-пронаторов. Эта информация полезна для клиницистов и специалистов по силовой и физической подготовке, которые работают со спортсменами, работающими над головой, поскольку эти упражнения могут быть назначены в качестве эффективного компонента программы упражнений по уходу за руками.
Авторы заявляют об отсутствии конфликта интересов.
1.
Вернер С.Л., Флейзиг Г.С., Диллман С.Дж., Эндрюс М.Р. Биомеханика локтя при подаче бейсбольного мяча. J Orthop Sports Phys Ther . 1993;17(6):274-278. doi:10.2519/jospt.1993.17.6.274
Google Scholar2.
Fleisig GS, Andrews JR, Dillman CJ, Escamilla RF. Кинетика подачи бейсбола с учетом механизмов травм. Ам Джей Спорт Мед . 1995;23(2):233-239. doi:10.1177/036354659502300218
Google Scholar3.
Morrey BF, An KN. Суставно-связочный вклад в стабильность локтевого сустава. Am J Sports Med . 1983;11(5):315-319. doi:10.1177/036354658301100506
Google Scholar4.
Leland DP, Conte S, Flynn N, et al. Распространенность хирургии медиальной локтевой коллатеральной связки у 6135 нынешних профессиональных бейсболистов: обновление 2018 г. Orthop J Sports Med . 2019;7(9):2325967119871442.
Google Scholar 5.
Дэвидсон П.А., Пинк М., Перри Дж., Джоб Ф.В. Функциональная анатомия группы мышц-сгибателей-пронаторов по отношению к медиальной коллатеральной связке локтевого сустава. Am J Sports Med . 1995;23(2):245-250. doi:10.1177/036354659502300220
Google Scholar6.
Park MC, Ahmad CS. Динамический вклад массы сгибателей-пронаторов в стабильность вальгусной деформации локтевого сустава. J Хирургия суставов костей Am . 2004;86(10):2268-2274. doi:10.2106/00004623-200410000-00020
Google Scholar7.
Lin F, Kohli N, Perlmutter S, Lim D, Nuber GW, Makhsous M. Вклад мышц в стабильность вальгусной деформации локтевого сустава. J Плечевой локтевой хирург . 2007;16(6):795-802. doi:10.1016/j.jse.2007.03.024
Google Scholar8.
Udall JH, Fitzpatrick MJ, McGarry MH, Leba TB, Lee TQ. Влияние нагрузки мышц-сгибателей-пронаторов на вальгусную стабильность локтевого сустава с интактной, растянутой и резецированной медиальной локтевой коллатеральной связкой. J Плечевой локтевой хирург . 2009;18(5):773-778. doi:10.1016/j.jse.2009.03.008
J Плечевой локтевой хирург . 2009;18(5):773-778. doi:10.1016/j.jse.2009.03.008
9.
Gregory B, Nyland J. Медиальная травма локтя у молодых спортсменов, занимающихся метанием. Мышцы Связки Сухожилия J . 2013;3(2):91-100. doi:10.32098/mltj.02.2013.06
Google Scholar10.
Otoshi K, Kikuchi S, Shishido H, Konno S. Проксимальное происхождение мышц-сгибателей-пронаторов и их роль в динамической стабилизации локтевого сустава сустав: анатомическое исследование. Сур Радиол Анат . 2014;36(3):289-294. doi:10.1007/s00276-013-1168-3
Google Scholar11.
Tajika T, Oya N, Ichinose T, et al. Вклад мышц-сгибателей-пронаторов в стабильность вальгусной деформации локтевого сустава: ультразвуковой анализ у школьных питчеров с симптомами и без них. JSES Int . 2020;4(1):9-14. doi:10.1016/j.jses.2019.10.003
Google Scholar PubMed Central PubMed12.
Millard N, DeMoss A, McIlvain G, Becket т Дж.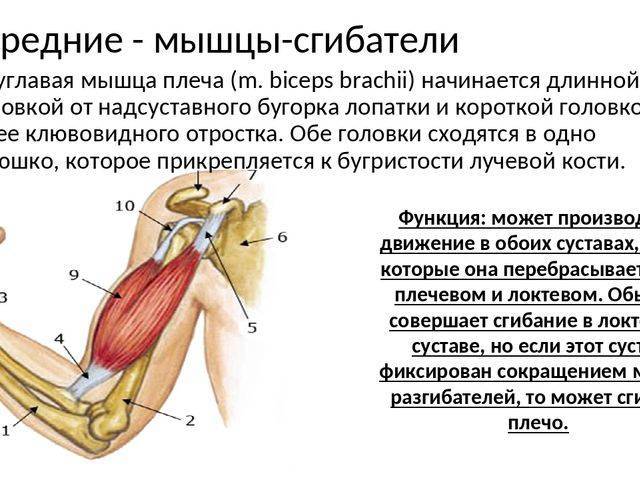 А., Яско Дж.Дж., Тиммонс М.К. Упражнение на сгибание запястья увеличивает ширину медиальной щели локтевого сустава во время вальгусного стресс-теста. J УЗИ Мед . 2019;38(4):959-966. doi:10.1002/jum.14779
А., Яско Дж.Дж., Тиммонс М.К. Упражнение на сгибание запястья увеличивает ширину медиальной щели локтевого сустава во время вальгусного стресс-теста. J УЗИ Мед . 2019;38(4):959-966. doi:10.1002/jum.14779
13.
Sisto DJ, Jobe FW, Moynes DR, Antonelli DJ. Электромиографический анализ локтя при подаче. Am J Sports Med . 1987;15(3):260-263. doi:10.1177/036354658701500314
Google Scholar14.
Digiovine NM, Jobe FW, Pink M, Perry J. Электромиографический анализ верхней конечности при качке. J Плечевой локтевой хирург . 1992;1(1):15-25. doi:10.1016/s1058-2746(09)80011-6
Google Scholar15.
Hamilton CD, Glousman RE, Jobe FW, Brault J, Pink M, Perry J. Динамическая стабильность локтевого сустава: электромиографический анализ группа сгибателей-пронаторов и группа разгибателей у кувшинов с вальгусной нестабильностью. J Плечевой локтевой хирург . 1996;5(5):347-354. doi:10.1016/s1058-2746(96)80065-6
Google Scholar 16.
Lipinski CL, Donovan L, McLoughlin TJ, Armstrong CW, Norte GE. Поверхностная электромиография мускулатуры предплечья во время восстановительной программы бросков через голову. Физ Тер Спорт . 2018;33:109-116. doi:10.1016/j.ptsp.2018.07.006
Google Scholar17.
Hirashima M, Kadota H, Sakurai S, Kudo K, Ohtsuki T. Последовательная мышечная активность и ее функциональная роль в верхней конечности и туловище во время метание через руку. J Sports Sci . 2002;20(4):301-310. doi:10.1080/026404102753576071
Google Scholar18.
Кили Д.В., Вике Дж., Алфорд К., Оливер Г.Д. Биомеханический анализ пронации предплечья и ее взаимосвязь с движением мяча для фастбола с двумя и четырьмя швами. Дж Прочность Сопротивление . 2010;24(9):2366-2371. doi:10.1519/jsc.0b013e3181b22aea
Google Scholar19.
Glousman RE, Barron J, Jobe FW, Perry J, Pink M. Электромиографический анализ локтевого сустава у здоровых и травмированных питчеров с недостаточностью медиальной коллатеральной связки . Am J Sports Med . 1992;20(3):311-317. doi:10.1177/036354659202000313
Am J Sports Med . 1992;20(3):311-317. doi:10.1177/036354659202000313
20.
Zipp P. Рекомендации по стандартизации положения отведений в поверхностной электромиографии. Europ J Appl Physiol . 1982;50(1):41-54. doi:10.1007/bf00952243
Google Scholar21.
Remaley DT, Fincham B, McCullough B, et al. Поверхностная электромиография мускулатуры предплечья во время игры в софтбол с ветряной мельницей. Orthop J Sports Med . 2015;3(1):232596711456679. doi:10.1177/2325967114566796
Google Scholar PubMed Central PubMed22.
Kendall FP, Kendall McCreary E, Provance PG , Роджерс М., Романи В. Мышцы: тестирование и функция, осанка и боль . Уолтерс Клувер Хелс; 2014.
Google Scholar23.
Андерсен Л.Л., Андерсен С.Х., Мортенсен О.С., Поулсен О.М., Бьорнлунд И.Б., Зебис М.К. Мышечная активация и воспринимаемая нагрузка во время реабилитационных упражнений: сравнение гантелей и эластичного сопротивления.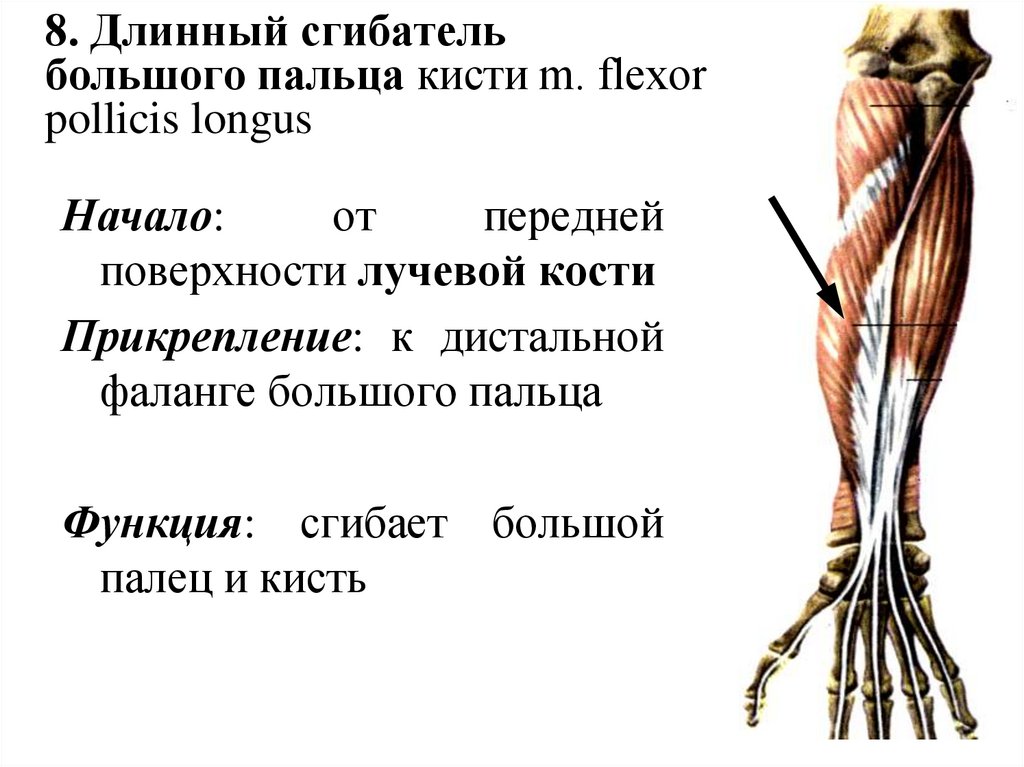 Физ Тер . 2010;90(4):538-549. doi:10.2522/ptj.200
Физ Тер . 2010;90(4):538-549. doi:10.2522/ptj.200
24.
Mullaney MJ, Perkinson C, Kremenic I, Tyler TF, Orishimo K, Johnson C. ЭМГ мышц плеча во время реактивных изометрических упражнений на упругое сопротивление. Int J Sports Phys Ther . 2017;12(3):417-424.
Google Scholar25.
Маллани М., Николас С., Тайлер Т., Фукунага Т., МакХью М. Уход за руками, основанный на доказательствах: еще раз о метателях 10. Int J Sports Phys Ther . 2021;16(6):1379-1386. doi:10.26603/001c.29900
Google Scholar PubMed Central PubMed26.
Orishimo KF, McHugh MP. Влияние эксцентрически смещенной домашней программы по укреплению подколенного сухожилия на силу сгибателей колена и соотношение длины и напряжения. Дж Прочность Сопротивление . 2015;29(3):772-778. doi:10.1519/jsc.0000000000000666
Google ScholarРастяжение сгибателей предплечья — разрыв
«Остеопатия основана на совершенстве работы природы. Когда все части человеческого тела в порядке, мы имеем здоровье. Когда их нет, следствием является болезнь. Когда части приспосабливаются, болезнь уступает место здоровью. Работа остеопата состоит в том, чтобы приспособить тело от ненормального к нормальному; затем ненормальное состояние уступает место нормальному, и здоровье является результатом нормального состояния». Доктор А. Т. Стилл
Когда все части человеческого тела в порядке, мы имеем здоровье. Когда их нет, следствием является болезнь. Когда части приспосабливаются, болезнь уступает место здоровью. Работа остеопата состоит в том, чтобы приспособить тело от ненормального к нормальному; затем ненормальное состояние уступает место нормальному, и здоровье является результатом нормального состояния». Доктор А. Т. Стилл
Предплечья являются неотъемлемой частью движения кистей и рук, поэтому боль в этой области может сильно мешать повседневной жизни. Мышцы-сгибатели/сухожилия — это мышцы, которые позволяют вам сгибать пальцы через сухожилия. Мышцы-сгибатели начинаются в локте и предплечье и переходят в сухожилия сразу за серединой предплечья. Затем они прикрепляются к костям пальцев.
Наиболее распространенные симптомы боли в предплечье включают слабость или потерю сцепления, боль и болезненность в мышцах между запястьем и локтем. Обычно при захвате чего-либо и во время активности, особенно движений рук. Мышцы предплечья ощущаются скованными, может быть некоторый отек. Сильные деформации связаны с большей болью и потерей силы. Вы также можете почувствовать острую, колющую боль, если попытаетесь согнуть запястье или надавить рукой на какой-либо предмет. Может быть чрезвычайно трудно согнуть руку вверх в локтевом суставе или полностью разогнуть руку с порванным или поврежденным разрывом или травмой сухожилия массы мышц-сгибателей. Мышечный спазм, при котором ваше предплечье напрягается без сгибания, является еще одним потенциально болезненным симптомом разрыва мышц предплечья.
Мышцы предплечья ощущаются скованными, может быть некоторый отек. Сильные деформации связаны с большей болью и потерей силы. Вы также можете почувствовать острую, колющую боль, если попытаетесь согнуть запястье или надавить рукой на какой-либо предмет. Может быть чрезвычайно трудно согнуть руку вверх в локтевом суставе или полностью разогнуть руку с порванным или поврежденным разрывом или травмой сухожилия массы мышц-сгибателей. Мышечный спазм, при котором ваше предплечье напрягается без сгибания, является еще одним потенциально болезненным симптомом разрыва мышц предплечья.
Растяжение мышц предплечья — это частичный или полный разрыв тонких волокон мышц предплечья. Мышцы предплечья позволяют разгибать и сгибать запястье и пальцы. Растяжение предплечья оценивается по шкале от 1 до 3. Степень 1 является легкой и не сопровождается потерей силы. Степень 2 включает более сильную боль и потерю силы. 3 степень — разрыв мышечно-сухожильного аппарата, требующий хирургического вмешательства.
Растяжение мышц предплечья может быть вызвано: хроническим перенапряжением, плохим биомеханическим движением или внезапной чрезмерной перегрузкой при подъеме тяжестей, неправильными повторяющимися спортивными приемами, длительной работой за компьютером, плохой эргономикой/осанкой при работе или ручным трудом/сделай сам, предыдущим напряжением или травмой в зону, внезапно перегружая мышцы предплечья, когда они не готовы к нагрузке, получая прямой удар по мышцам предплечья. Чрезмерное хватание и повторяющиеся движения могут привести к нагрузке на предплечья: удерживание теннисной ракетки, хватание за ручки велосипеда, захват небольшого инструмента, такого как отвертка, удерживание и перемещение кисти, перенос продуктов в сумке и т. д. Может возникнуть боль. постепенно (как хроническая проблема) или внезапно из-за резкого движения (как острая травма). В течение недель и месяцев травма становится болезненной и ограниченной.
Фасция — это соединительная ткань, которая, словно паутина, обвивается вокруг мышц и реагирует на то, как вы используете свое тело. Плохие модели движений могут привести к постепенному стягиванию фасций в спайки, что не позволит мышцам полностью расслабиться и удлиниться. Когда это происходит, мышца может прилипнуть к соседней мышце, заставляя их работать усерднее и вызывая напряжение. Со временем спайки становятся более распространенными. Это «торможение» движения мышц создаст ощущение, что ваши мышцы ослабли, они не могут выполнять столько работы, сколько раньше, и боль становится обычным явлением.
Плохие модели движений могут привести к постепенному стягиванию фасций в спайки, что не позволит мышцам полностью расслабиться и удлиниться. Когда это происходит, мышца может прилипнуть к соседней мышце, заставляя их работать усерднее и вызывая напряжение. Со временем спайки становятся более распространенными. Это «торможение» движения мышц создаст ощущение, что ваши мышцы ослабли, они не могут выполнять столько работы, сколько раньше, и боль становится обычным явлением.
Разрыв предплечья может серьезно помешать вам работать на компьютере или поддерживать физическую форму. Большинство травм предплечья могут быть диагностированы остеопатом путем сбора анамнеза и простого физического осмотра. Правильный диагноз приведет к более быстрому выздоровлению. Остеопат также объяснит возможные факторы, которые могли способствовать травме, такие как снижение подвижности запястья, локтя или плеча, плохая осанка или движения и неподходящие упражнения. Остеопатическое лечение может включать миофасциальное расслабление, артикуляцию суставов или более тонкие краниальные техники.
