2 Кости голени и стопы, их соединения. Пассивные и активные «затяжки» сводов стопы, механизм их действия на стопу.9(II) Кости голени и стопы, их соединения
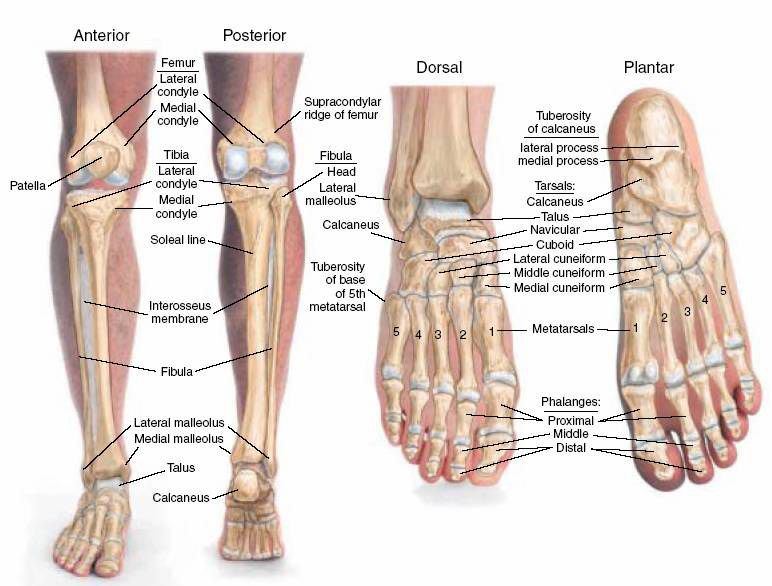
Большеберцовая кость на проксимальном эпифизе имеет:
медиальный и латеральный мыщелки с суставными поверхностями;
межмыщелковое возвышение с медиальным и латеральным бугорками:, передним и задним полем;
малоберцовую суставную поверхность с латеральной стороны ниже мыщелка.
На диафизе большеберцовой кости располагаются:
передний край (острый) – кверху переходит в бугристость, латеральный край, обращенный к малоберцовой кости и медиальный край;
поверхности: медиальная, латеральная и задняя с линией камбаловидной мышцы.
Дистальный эпифиз имеет:
малоберцовую вырезку по латеральному краю;
медиальную лодыжку с лодыжечной бороздой сзади;
суставные
поверхности: лодыжечную и нижнюю.
Ядра окостенения в большеберцовой кости появляются в проксимальном эпифизе в конце плодного периода, в дистальном – на 2-м году жизни, в диафизе – в начале плодного периода. Окончательное окостенение происходит в 19-24 года, лодыжки – в 16-19 лет.
Малоберцовая кость имеет:
на проксимальном эпифизе головку, шейку, на головке – верхушку и суставную поверхность;
на диафизе: передний, задний и межкостный края; латеральную, заднюю и медиальную поверхности;
на дистальном эпифизе: латеральную лодыжку с суставной поверхностью и ямкой (сзади).
Вторичные ядра окостенения появляются в дистальном эпифизе на 2-м году жизни, в проксимальном – на 3-5 годам, окончательное окостенение – к 20-24 годам.
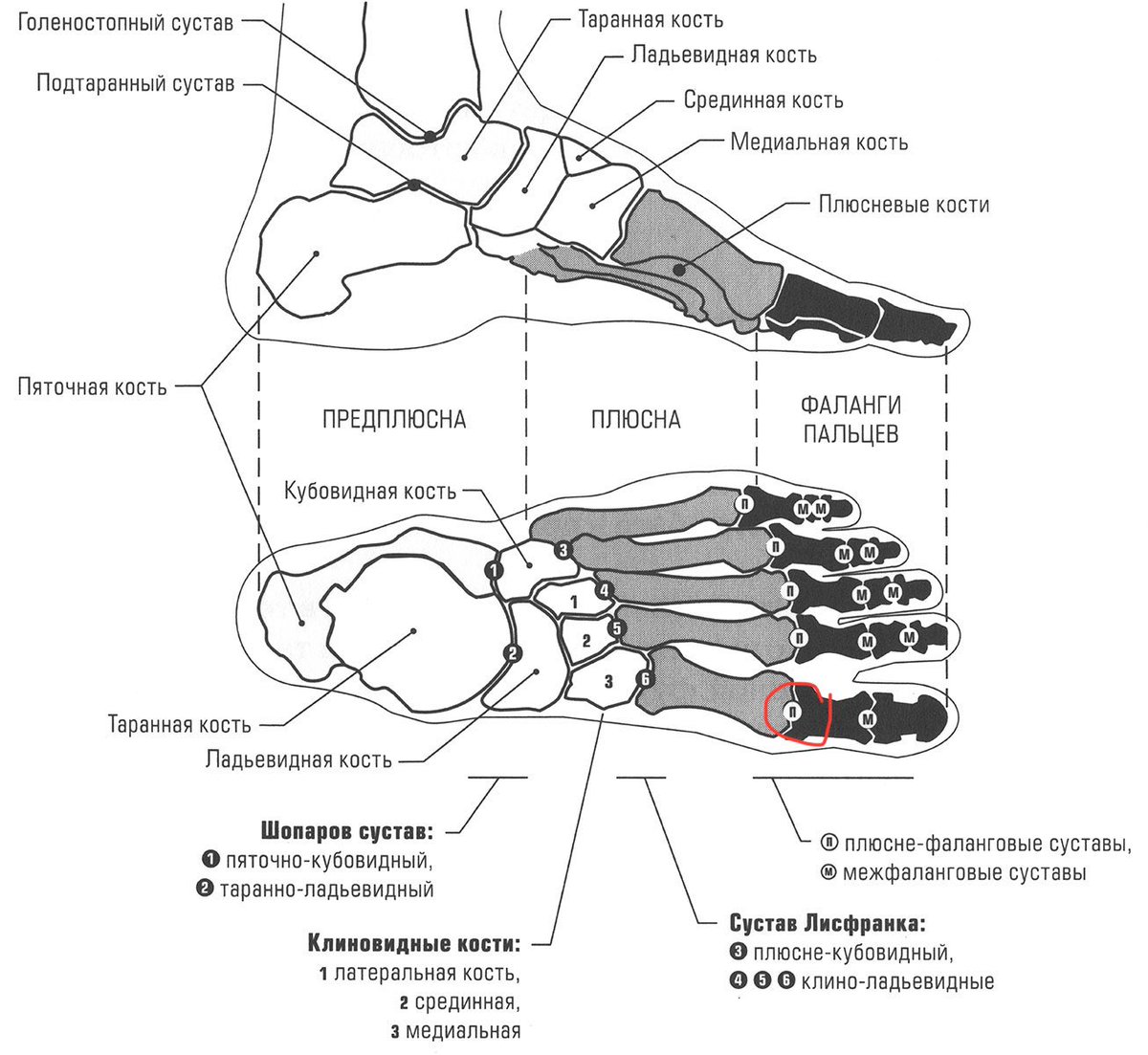
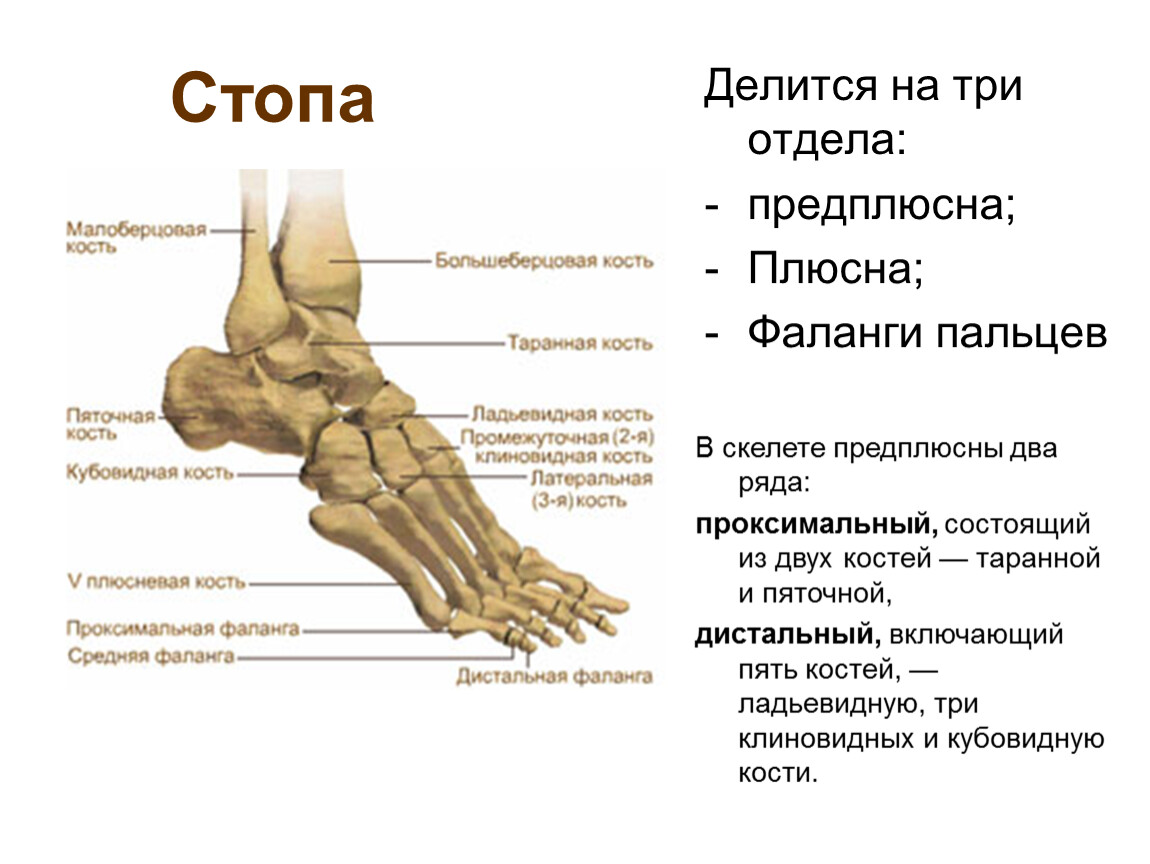
Кости
стопы подразделяются на кости предплюсны:
7 коротких губчатых костей и кости плюсны
в составе 5 коротких трубчатых костей;
а также фаланги пальцев – по три в каждом
пальце, кроме первого, имеющего две
фаланги.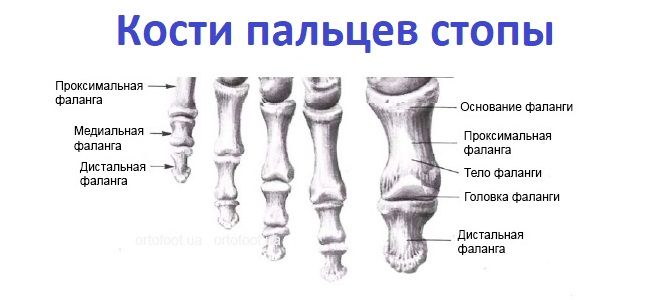
Таранная кость состоит из тела, головки, шейки. Сверху на теле находится блок с верхней, медиальной и латеральной лодыжечными суставными поверхностями. Снизу на теле располагаются пяточные суставные поверхности: передняя, средняя и задняя. Между средней и задней пяточными поверхностями находится борозда таранной кости. На головке лежит ладьевидная суставная поверхность. Таранная кость имеет отростки: латеральный и задний, на последнем – медиальный и латеральный бугорки, разделенные бороздой для сухожилия длинного сгибателя большого пальца.
Пяточная
кость имеет тело, заканчивающееся кзади
пяточным бугром. На теле различают
таранные суставные поверхности: переднюю,
среднюю и заднюю. Между средней и задней
находится борозда пяточной кости,
которая вместе с бороздой таранной
кости образует пазуху предплюсны.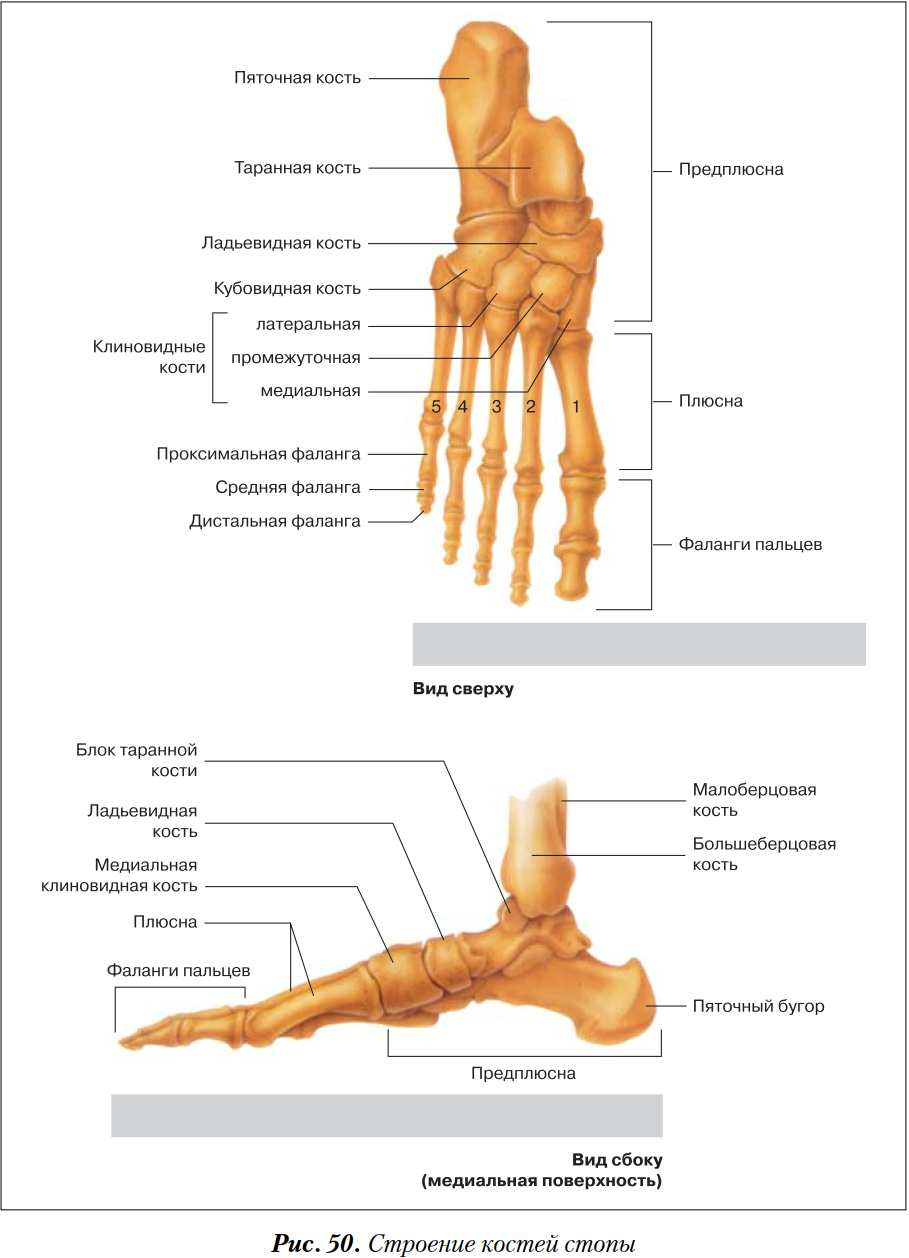
Ладьевидная кость на дистальном конце располагает тремя суставными поверхностями для клиновидных костей, бугристостью по медиальному краю для прикрепления задней большеберцовой мышцы.
Клиновидные кости имеют характерную треугольную форму, а на передней и задней сторонах и по бокам – суставные поверхности – спереди для сочленения с первыми тремя плюсневыми костями, сзади для соединения с ладьевидной костью, по бокам для сочленения между собой и кубовидной костью.
Кубовидная
кость тоже располагает суставными
поверхностями сзади для сочленения с
пяточной, спереди – с пястными костями,
а по медиальной поверхности – для
соединения латеральной клиновидной и
ладьевидной костями. На нижней поверхности
находится бугристость и борозда для
сухожилия длинной малоберцовой мышцы.
Кости плюсны и фаланги пальцев – это короткие трубчатые кости, каждая из которых состоит из основания, тела и головки. Суставные поверхности находятся на головке и основании. Головка первой плюсневой кости снизу разделена на две площадки, с которыми соприкасаются сесамовидные кости. На латеральной поверхности Y плюсневой кости находится бугристость для прикрепления короткой малоберцовой мышцы. Каждая дистальная (ногтевая) фаланга заканчивается бугорком для прикрепления мышечных сухожилий.
Первичные ядра окостенения появляются в пяточной кости на 6-м месяце плодного периода, в таранной – на 7-8, в кубовидной – на 9-м месяце. В остальных костях предплюсны формируются вторичные ядра: в латеральной клиновидной – на 1-м году, в медиальной клиновидной – на 3-4, в ладьевидной – на 4-5 году, полное окостенение – в 12-16 лет.
Плюсневые
кости начинают окостенение в эпифизах
в 3-6 лет, полное окостенение – в 12-16 лет.
Диафизы фаланг и плюсневых костей
первичные ядра формируют в 12-14 недель
плодного периода, вторичные – в фалангах
– в 3-4 года, полное окостенение фаланг
– в 18-20 лет.
Кости голени имеют соединения:
межберцовый сустав, образованный плоскими суставными поверхностями малоберцовой головки и верхнего эпифиза (боковая часть), большеберцовой кости; суставная капсула туго натянута и укреплена передней и задней связками головки малоберцовой кости; объем движений ограниченный; иногда межберцовый сустав может сообщаться с полостью коленного сустава;
межберцовый синдесмоз – непрерывное соединение короткими и толстыми фиброзными волокнами между малоберцовой вырезкой на дистальном эпифизе большеберцовой кости и суставной поверхностью латеральной лодыжки, укрепленное передней и задней межберцовыми связками, в межберцовый синдесмоз впячивается синовиальная мембрана голеностопного сустава;
межкостную
перепонку голени – фиброзную мембрану,
натянутую между межкостными краями
больше- и малоберцовой костей в области
их диафизов; она имеет вверху и внизу
отверстия для прохождения сосудов и
нервов.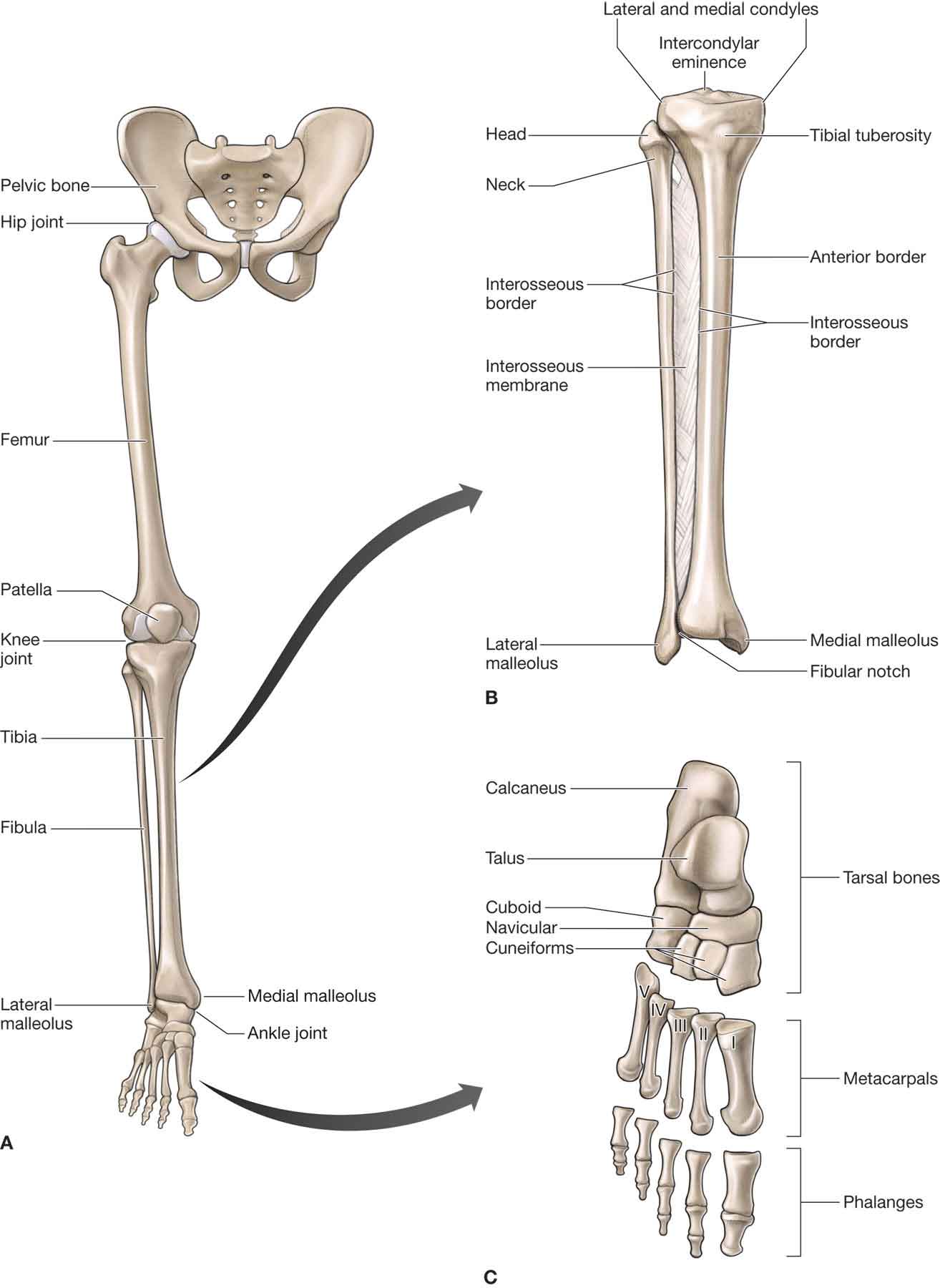
В соединениях костей голени движения почти отсутствуют, что связано с опорной функцией нижней конечности и формированием подвижного голеностопного сустава.
В образовании голеностопного сустава участвуют:
большеберцовая кость своей нижней и медиальной лодыжечной суставными поверхностями;
малоберцовая кость — суставной поверхностью латеральной лодыжки;
таранная кость — верхней и лодыжечными (медиальной и латеральной суставными поверхностями), расположенными на блоке.
Лодыжки вилкообразно охватывают блок, образуя типичный блоковидный сустав, сложный из-за сочленения трех костей.
Суставная капсула сзади прикрепляется по краю суставных поверхностей сочленяющихся костей, спереди на 0,5-1 см выше ее.
По бокам капсула толстая и прочная, спереди и сзади – тонкая, рыхлая, складчатая, усилена мощными боковыми связками:
медиальной (дельтовидной) – толстой, прочной, веером расходящейся от медиальной лодыжки к таранной, ладьевидной и пяточной костям; в связке выделяют части: большеберцово-ладьевидную, большеберцово-пяточную, переднюю и заднюю большеберцово-таранные;
латеральной
в составе передней таранно-малоберцовой,
задней таранно-малоберцовой,
пяточно-малоберцовой связок.
Связки настолько прочны, особенно медиальная, что при максимальном растяжении способны оторвать лодыжку.
В голеностопном суставе выполняются движения:
сгибание (подошвенное) и разгибание (тыльное сгибание) вокруг фронтальной оси с размахом в 60 о;
приведение и отведение вокруг сагиттальной оси с размахом в 17 о;
пронация и супинация вокруг вертикальной оси с размахом в 22 о.
Подтаранный сустав образован задними суставными поверхностями таранной и пяточной костей, которые хорошо соответствуют друг другу. Капсула у сустава тонкая, натянутая, укреплена голеностопными связками.
Таранно-пяточно-ладьевидный
сустав образован суставной поверхностью
на головке талуса, которая спереди
сочленяется с ладьевидной костью, а
снизу – с пяточной. Суставная поверхность
пяточной кости дополняется подошвенной
пяточно-ладьевидной связкой, которая
у головки таранной кости переходит в
волокнистый хрящ. Суставная капсула
прикрепляется по краю суставных
поверхностей, замыкая единую полость.
Капсула укрепляется прочными связками:
межкостной таранно-пяточной до 0,5 см
толщиной и сверху — таранно-ладьевидной.
При растяжении их (особенно таранно-пяточной
связки) опускается головка талуса и
стопа уплощается.
Суставная капсула
прикрепляется по краю суставных
поверхностей, замыкая единую полость.
Капсула укрепляется прочными связками:
межкостной таранно-пяточной до 0,5 см
толщиной и сверху — таранно-ладьевидной.
При растяжении их (особенно таранно-пяточной
связки) опускается головка талуса и
стопа уплощается.
По форме сустав шаровидный, но с одной сагиттальной осью движения. Вместе с подтаранным составляет комбинированный сустав с ограниченным объемом движений в виде приведения и отведения, поворота стопы кнаружи и во внутрь. Общий объем всех движений не превышает 55 о. У новорожденного и грудного ребенка данный сустав находится в супинированном положении, с началом ходьбы постепенно происходит пронирование с опусканием медиального края стопы.
Пяточно-кубовидный
сустав – седловидный с большой
конгруэнтностью суставных поверхностей;
суставная капсула с медиальной стороны
толстая и натянутая, с латеральной –
тонкая и свободная, укреплена связками:
подошвенной пяточно-кубовидной и длинной
подошвенной. Движения в ограниченном
объеме осуществляются вокруг продольной
оси в виде небольшого вращения,
дополняющего движения в
таранно-пяточно-ладьевидном суставе.
Движения в ограниченном
объеме осуществляются вокруг продольной
оси в виде небольшого вращения,
дополняющего движения в
таранно-пяточно-ладьевидном суставе.
Поперечный (Шопаров) сустав предплюсны включает пяточно-кубовидный и таранно-ладьевидный суставы так, что суставная щель приобретает вид латинского S, проходящего поперек стопы. Общая связка суставов – раздвоенная (lig. bifurcatum) — служит своеобразным ключом, при рассечении которого широко открывается полость сустава. Она имеет Y-образный вид и состоит из пяточно-ладьевидной и пяточно-кубовидной связок.
Клиновидно-ладьевидный сустав – плоский, сложный, так как образуется тремя клиновидными и одной ладьевидной костями, укреплен тыльными и подошвенными клино-ладьевидными и внутрисуставными межклиновидными связками. Полость сустава может сообщаться с предплюсне-плюсневыми суставами.
Предплюсне-плюсневые (Лисфранковы) суставы – плоские с ограниченным объемом движений, включают следующие три сочленения:
сустав между медиальной клиновидной и первой плюсневой костями;
сустав между промежуточной, латеральной клиновидными костями и II, III плюсневыми;
сустав между кубовидной костью и IY, Y плюсневыми костями;
предплюсне-плюсневые
суставы укреплены тарзо-метатарзальными
связками (дорсальными и плантарными) и
межкостными клиноплюсневыми связками,
из которых медиальная клиноплюсневая
связка считается в практике ключом этих
суставов.
Межплюсневые суставы – плоские, образованы боковыми поверхностями оснований плюсневых костей, укреплены тыльными и подошвенными плюсневыми межкостными связками. Движения в них сильно ограничены.
Плюснефаланговые суставы образованы шаровидными головками плюсневых костей и вогнутыми основаниями проксимальных фаланг. Капсулы имеют тонкие, свободные, укрепленные коллатеральными и подошвенной связками. Глубокая поперечная плюсневая связка соединяет между собой головки плюсневых костей и укрепляет капсулы суставов. Сгибание и разгибание в них – 90 о, приведение и отведение – ограничено.
Межфаланговые суставы – проксимальные и дистальные, кроме сустава большого пальца, где один межфаланговый сустав. Они относятся к блоковидным суставам, укреплены коллатеральными и подошвенными связками.
Стопа
человека утратила приспособления
хватательного органа, но приобрела
своды, необходимые для опоры и передвижения
всего тела.
Кости предплюсны и плюсневые соединяются между собой малоподвижными суставами.
Причем, за счет формы костей и их «тугих» сочленений образовались выпуклые кверху пять продольных и один поперечный своды с точками опоры на пяточный бугор и головки I и Y плюсневых костей.
Продольные своды проходят по длинной оси плюсневых костей: от их головок, через тело к основанию и далее через прилежащие кости предплюсны до пяточного бугра, где все они сходятся. Наиболее длинным и высоким является второй свод.
Медиальные продольные своды выполняют рессорную (пружинящую) функцию, латеральные – опорную. На уровне наиболее высоких точек продольных сводов проходит свод поперечный.
Своды
удерживаются благодаря форме костей и
суставов, укрепленных пассивными
«затяжками» — связками с подошвенным
апоневрозом и активными «затяжками» –
мышцами и их сухожилиями. Продольные
своды укрепляют следующие связки:
длинная подошвенная, пяточно-ладьевидная
и подошвенный апоневроз. Поперечный
свод удерживают метатарзальные поперечные
и межкостные связки. Сухожилия длинных
сгибателей и разгибателей пальцев,
большеберцовых мышц, а также короткие
сгибатели пальцев, червеобразные мышцы
укрепляют продольные своды. Поперечный
свод удерживается квадратной мышцей
подошвы, приводящими и межкостными
мышцами.
Поперечный
свод удерживают метатарзальные поперечные
и межкостные связки. Сухожилия длинных
сгибателей и разгибателей пальцев,
большеберцовых мышц, а также короткие
сгибатели пальцев, червеобразные мышцы
укрепляют продольные своды. Поперечный
свод удерживается квадратной мышцей
подошвы, приводящими и межкостными
мышцами.
В процессе эволюции человека в стопе произошли крупные изменения, связанные с ее сводчатым строением. На подошве образовалось три опорных пункта – пятка и основания первого и пятого пальцев, кости предплюсны увеличились в размерах, а фаланги пальцев значительно укоротились. Соединения стопы стали тугоподвижными. Прикрепление длинных мышц голени и коротких мышц стопы сместилось в медиальном направлении к большому пальцу, что усиливает пронацию стопы и укрепляет своды. Потеря приобретенных свойств сопровождается развитием плоскостопия, что для человека является болезнью.
Древние люди отличались ногами
4772
Добавить в закладки
Международная группа антропологов и палеонтологов под руководством Кэрол Вард (Carol Ward) из университета Миссури изучила, как отличались между собой ноги древних предков человека, сообщается на официальном сайте института. Оказалась, что по остаткам нижних конечностей, также как и по черепу, можно узнать, к какому виду принадлежал наш предок. Результаты исследования были опубликованы в свежем номере Journal of Human Evolution.
Отправной точкой их изысканий стал фрагмент кости ноги человека неизвестного вида, найденный в Кооби-Фора еще в 1980 году. В 2009 году при раскопках был найден ее другой фрагмент, и ученые получили в свое распоряжение почти целую правую бедренную кость древнего человека возрастом около 1,9 млн лет. Требовалось установить, чья это нога.
Кооби-Фора — территория на севере Кении, известная тем, что там
было найдено большое количество останков древних представителей
рода Homo. Данные из этой местности позволили ученым
получить сведения о том, как выглядели наши далекие предки.
Однако до сих пор внимание исследователей было сосредоточено,
прежде всего, на черепах и челюстях древних Homo,
которые, как известно, сильно отличались между собой у разных
видов. Но есть немало находок, когда от древнего человека
сохранились кости ног, рук и других частей тела, но не голова.
Определить по таким данным, что это за вид, практически не
представлялось возможным.
Данные из этой местности позволили ученым
получить сведения о том, как выглядели наши далекие предки.
Однако до сих пор внимание исследователей было сосредоточено,
прежде всего, на черепах и челюстях древних Homo,
которые, как известно, сильно отличались между собой у разных
видов. Но есть немало находок, когда от древнего человека
сохранились кости ног, рук и других частей тела, но не голова.
Определить по таким данным, что это за вид, практически не
представлялось возможным.
Группа Кэрол Вард попыталась справиться с этой проблемой. Для
этого группа Кэрол Вард сравнила ее с большим количеством других
костей ног, про владельцев которых ученые знали, к какому виду
они относились. Найденная нога по строению больше напоминает ногу Homo Erectus (человека умелого), однако самые ранние
найденные останки особей этого вида относятся к периоду примерно
1,8 млн лет назад. Поэтому, скорее всего, ученые имеют дело с
ногой более ранних Homo Habilis или Homo
Rudolfensis, хотя и не типичной для этих видов. Возможно,
это различие было связано с тем, что хозяин ноги вел какой-то
другой образ жизни по сравнению с типичными Homo
Habilis.
Возможно,
это различие было связано с тем, что хозяин ноги вел какой-то
другой образ жизни по сравнению с типичными Homo
Habilis.
Как отмечает Кэрол Вард, в ходе исследования оказалось, что строение нижних конечностей у древних людей отличались не в меньшей степени, чем их черепа, и соответственно могут служить для того, чтобы определить видовую принадлежность находки. Кости нижних конечностей различных видов отличаются, например, у Homo Erectus по сравнению с Homo Habilis более тонкий таз.
Научная Россия недавно рассказывала об еще одной находке костей предка человека из Восточной Африки.
homo habilis палеонтологические находки
Информация предоставлена Информационным агентством «Научная Россия». Свидетельство о регистрации СМИ: ИА № ФС77-62580, выдано Федеральной службой по надзору в сфере связи, информационных технологий и массовых коммуникаций 31 июля 2015 года.
Кость ноги животного — Etsy Турция
Etsy больше не поддерживает старые версии вашего веб-браузера, чтобы обеспечить безопасность пользовательских данных. Пожалуйста, обновите до последней версии.
Пожалуйста, обновите до последней версии.
Воспользуйтесь всеми преимуществами нашего сайта, включив JavaScript.
Найдите что-нибудь памятное, присоединяйтесь к сообществу, делающему добро.
( 106 релевантных результатов, с рекламой Продавцы, желающие расширить свой бизнес и привлечь больше заинтересованных покупателей, могут использовать рекламную платформу Etsy для продвижения своих товаров. Вы увидите результаты объявлений, основанные на таких факторах, как релевантность и сумма, которую продавцы платят за клик. Узнать больше. )Пилотное исследование потенциальной взаимосвязи между длиной костей голени и результатами спринта у спринтеров; есть ли какие-либо различия в беге на 100 и 400 м в зависимости от вида спорта? | BMC Research Notes
- Research note
- Открытый доступ
- Опубликовано:
- Даичи Томита 1 ,
- Тадаши Суга ORCID: orcid.org/0000-0002-0313-9660 1 ,
- Масафуми Терада 1 ,
- Такахиро Танака 1 , 900 13 Юто Мияке 1 ,
- Хиромаса Уэно 1,2, 3 ,
- Мицуо Оцука 1 ,
- Акинори Нагано 1 и
- …
- Тадао Исака 1
Исследовательские заметки BMC том 13 , Номер статьи: 297 (2020) Процитировать эту статью
6213 Доступ
3 Цитаты
2 Альтметрика
Сведения о показателях
Abstract
Объектив
В этом исследовании изучалась взаимосвязь между длиной костей голени и результатами спринта у спринтеров. Длина костей голени у 28 специализированных спринтеров на 100 м и 28 специализированных спринтеров на 400 м была измерена с помощью магнитно-резонансной томографии. Длины костей бедра и голени оценивали путем расчета длины бедренной и большеберцовой костей соответственно. Чтобы свести к минимуму различия в размерах тела среди участников, обе длины костей были нормализованы к росту. Отношение длины большеберцовой кости к длине бедренной кости рассчитывали для оценки взаимодействия между длинами костей верхней и нижней частей голени. В качестве параметров спринтерских результатов использовались баллы Международной любительской легкоатлетической федерации (ИААФ), основанные на личном лучшем времени спринтеров в каждой группе.
Длина костей голени у 28 специализированных спринтеров на 100 м и 28 специализированных спринтеров на 400 м была измерена с помощью магнитно-резонансной томографии. Длины костей бедра и голени оценивали путем расчета длины бедренной и большеберцовой костей соответственно. Чтобы свести к минимуму различия в размерах тела среди участников, обе длины костей были нормализованы к росту. Отношение длины большеберцовой кости к длине бедренной кости рассчитывали для оценки взаимодействия между длинами костей верхней и нижней частей голени. В качестве параметров спринтерских результатов использовались баллы Международной любительской легкоатлетической федерации (ИААФ), основанные на личном лучшем времени спринтеров в каждой группе.
Результаты
Значимой корреляции между абсолютной и относительной длиной бедренной и большеберцовой костей и баллами ИААФ у спринтеров на 100 и 400 м обнаружено не было. Напротив, отношение длины большеберцовой кости к длине бедра достоверно коррелировало с оценкой ИААФ у спринтеров на 400 м ( r = 0,445, P = 0,018), но не у спринтеров на 100 м. Эти данные свидетельствуют о том, что длина костей голени может играть важную роль в достижении превосходных результатов в длинном спринте у специализированных спринтеров на дистанции 400 м.
Эти данные свидетельствуют о том, что длина костей голени может играть важную роль в достижении превосходных результатов в длинном спринте у специализированных спринтеров на дистанции 400 м.
Введение
Некоторые морфологические факторы связаны с превосходными результатами спринтеров у спринтеров [1,2,3,4,5,6,7]. Скорость спринта выражается как произведение длины и частоты шагов [8], что позволяет предположить, что морфологические факторы, регулирующие две спринтерские переменные, могут играть важную роль в достижении превосходных результатов в спринте у спринтеров. Длина ноги является основным морфологическим фактором, регулирующим длину шага [9, 10]; таким образом, более длинная нога может привести к лучшим результатам в спринте из-за увеличения длины шага во время спринта [11]. Мы и другие ранее обнаружили положительную связь между длиной ног и беговыми характеристиками у бегунов на выносливость [12,13,14]. Однако влияние длины ног на спринтерские результаты у спринтеров до сих пор плохо изучено.
Предыдущие исследования показали, что частота шагов может быть более важным кинематическим фактором для достижения превосходных результатов в спринте, чем длина шагов [15,16,17,18]. Морин и др. [15] продемонстрировали положительную корреляцию между частотой шагов и скоростью бега на 100 м; однако такой корреляции с длиной шага получено не было. Хобара и др. [16] сообщили, что частота шагов коррелирует с вертикальной жесткостью (то есть отношением пиковой вертикальной силы и вертикального смещения центра масс), которая положительно связана со скоростью спринта во время бега на 400 м. Однако такой корреляции не было получено и с длиной шага. Следовательно, морфологические факторы, регулирующие частоту шагов, могут быть более важными для достижения превосходных результатов в спринте, чем регулирующие длину шага.
Известно, что физиологические факторы, в том числе анаэробная способность и сопротивление утомлению, играют важную роль в достижении превосходных результатов в беге на 400 м [19, 20]. Использование этих физиологических возможностей во время спринта на 400 м может быть смягчено за счет экономичного спринта. Нога качается вперед с согнутым коленом во время фазы маха во время спринта. Основываясь на таком поведении ног, увеличение отношения длины голени к длине бедра (т. е. длины бедра) может уменьшить момент инерции ноги и положительную работу, выполняемую сгибателями бедра во время фазы переноса, возможно, из-за уменьшения масса ноги, так как масса голени меньше, чем масса бедра [21]. Следовательно, эта благоприятная морфология может быть полезна для увеличения частоты шагов и выполнения экономичного спринта. Принимая во внимание эти результаты, мы предположили, что более высокое отношение длины голени к длине бедра будет положительно связано с лучшими результатами в спринте у спринтеров, особенно у специализированных спринтеров на 400 м.
Использование этих физиологических возможностей во время спринта на 400 м может быть смягчено за счет экономичного спринта. Нога качается вперед с согнутым коленом во время фазы маха во время спринта. Основываясь на таком поведении ног, увеличение отношения длины голени к длине бедра (т. е. длины бедра) может уменьшить момент инерции ноги и положительную работу, выполняемую сгибателями бедра во время фазы переноса, возможно, из-за уменьшения масса ноги, так как масса голени меньше, чем масса бедра [21]. Следовательно, эта благоприятная морфология может быть полезна для увеличения частоты шагов и выполнения экономичного спринта. Принимая во внимание эти результаты, мы предположили, что более высокое отношение длины голени к длине бедра будет положительно связано с лучшими результатами в спринте у спринтеров, особенно у специализированных спринтеров на 400 м.
Обычно длину ноги измеряют антропометрическим методом, который выполняется вручную с помощью рулетки. Тем не менее, это измерение может иметь техническое ограничение из-за возможной неоднозначности измерения длины ноги. По сравнению с этим общим антропометрическим измерением магнитно-резонансная томография (МРТ) больше подходит для морфологических измерений [1, 2, 3, 4, 5, 6, 7, 14, 22], включая измерения длины костей [4, 5, 14, 22]. 22]. Чтобы проверить нашу гипотезу с большей точностью, мы использовали МРТ для изучения взаимосвязи между переменными длины кости ноги и результатами спринта у специализированных спринтеров на 100 и 400 м.
По сравнению с этим общим антропометрическим измерением магнитно-резонансная томография (МРТ) больше подходит для морфологических измерений [1, 2, 3, 4, 5, 6, 7, 14, 22], включая измерения длины костей [4, 5, 14, 22]. 22]. Чтобы проверить нашу гипотезу с большей точностью, мы использовали МРТ для изучения взаимосвязи между переменными длины кости ноги и результатами спринта у специализированных спринтеров на 100 и 400 м.
Основной текст
Методы
Испытуемые
В этом исследовании приняли участие 56 мужчин-спринтеров. Из них 28 спринтеров (возраст: 20,3 ± 1,6 года) специализировались в беге на 100 м и 28 спринтеров (возраст: 20,1 ± 1,4 года) специализировались на беге на 400 м. Спринтеры в каждой группе были хорошо подготовлены и регулярно соревновались на своих дистанциях. Среднее личное лучшее время бегунов-специалистов на 100 м составило 11,05 ± 0,33 с. Среднее личное лучшее время специализированных спринтеров на 400 м составило 490,96 ± 1,63 с. Средние баллы Международной любительской легкоатлетической федерации (межиндивидуальные баллы для сравнения соревновательных результатов в различных видах спорта), основанные на этом личном лучшем времени, использовались в качестве параметров спринтерских результатов в каждой группе. Информированное письменное согласие было получено от всех участников. Это исследование было одобрено Комитетом по этике Университета Рицумейкан.
Информированное письменное согласие было получено от всех участников. Это исследование было одобрено Комитетом по этике Университета Рицумейкан.
МРТ-измерения
Репрезентативные изображения для расчета длины костей голени с помощью магнитно-резонансной томографии (МРТ) показаны на рис. 1. МРТ-измерение было описано ранее [14]. Анализы для измерения длины костей ног проводились с использованием программного обеспечения для анализа изображений (OsiriX Version 5.6; OsiriX Foundation, Женева, Швейцария). Длины бедренной и большеберцовой костей были рассчитаны по коронарным изображениям ноги соответственно. Длину бедренной кости рассчитывали как расстояние между верхушкой большого вертела и дистальным концом латерального мыщелка бедренной кости. Длину большеберцовой кости рассчитывали как расстояние между проксимальным концом латерального мыщелка и дистальной нижней поверхностью большеберцовой кости. Общая длина бедренной и большеберцовой костей рассчитывалась для оценки общей длины костей голени.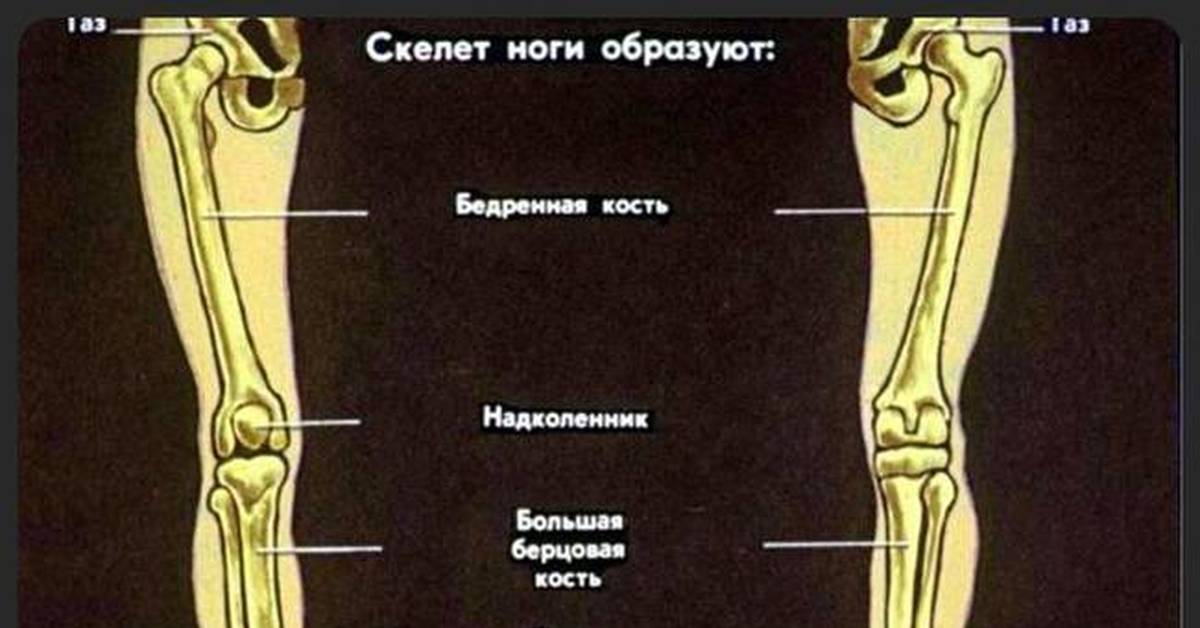 Чтобы свести к минимуму различия в размерах тела среди участников, эти обе длины были нормализованы к росту. Отношение длины большеберцовой кости к длине бедренной кости рассчитывали для оценки взаимодействия между длинами костей верхней и нижней частей голени. Воспроизводимость длины кости голени была отмечена в нашем предыдущем исследовании [14].
Чтобы свести к минимуму различия в размерах тела среди участников, эти обе длины были нормализованы к росту. Отношение длины большеберцовой кости к длине бедренной кости рассчитывали для оценки взаимодействия между длинами костей верхней и нижней частей голени. Воспроизводимость длины кости голени была отмечена в нашем предыдущем исследовании [14].
Репрезентативные снимки магнитно-резонансной томографии, используемые для измерения длины бедренной и большеберцовой костей. Длину бедренной кости измеряли как расстояние между верхушкой большого вертела и дистальным концом латерального мыщелка бедренной кости. Длину большеберцовой кости измеряли как расстояние между проксимальным концом латерального мыщелка и дистальной нижней поверхностью большеберцовой кости. Сравнение групп проводили с использованием непарного 9-ти0186 т -тест. Связь между переменными оценивалась с использованием корреляции моментов произведения Пирсона. Статистическая значимость была определена как P <0,05. Все статистические анализы проводились с использованием программного обеспечения IBM SPSS (версия 19.0; International Business Machines Corp, Нью-Йорк, США).
Все статистические анализы проводились с использованием программного обеспечения IBM SPSS (версия 19.0; International Business Machines Corp, Нью-Йорк, США).
Результаты
Показатели ИААФ существенно не отличались между специализированными спринтерами на 100 м и на 400 м (873 ± 97 и 863 ± 97).
Физические характеристики и переменные длины кости голени бегунов на 100 и 400 м приведены в таблице 1. Физические характеристики (т. е. рост, масса тела и индекс массы тела) не различались между 100 и 400 м. -м спринтеры. Кроме того, все переменные длины кости ноги не отличались между двумя группами.
Таблица 1 Физические характеристики и переменные длины костей голени у специализированных спринтеров на 100 и 400 м в Таблице 2. Абсолютная и относительная длина бедренной и большеберцовой костей не коррелировала с оценками ИААФ как у спринтеров на 100 м, так и на 400 м. Абсолютная и относительная общая длина бедренной и большеберцовой костей также не коррелировала с баллом IAAF в двух группах. Напротив, отношение длины большеберцовой кости к длине бедренной кости коррелирует с оценкой ИААФ у спринтеров на 400 м, но не у спринтеров на 100 м. Таблица 2 Коэффициенты корреляции между переменными длины костей голени и результативностью в спринте (оценка Международной любительской легкоатлетической федерации) у специализированных спринтеров на 100 и 400 м более высокое отношение длины голени к длине бедренной кости коррелировало с лучшим результатом ИААФ у спринтеров на 400 м, но не у спринтеров на 100 м. Поддержание постоянной частоты шагов во время бега на 400 м необходимо для достижения превосходных результатов в длинном спринте [16, 23]. Морфологические факторы могут способствовать поддержанию частоты шагов, потенциально поддерживая экономичный спринт [5, 6]. Соотношение между длинами костей голени может быть полезным для уменьшения момента инерции ноги и положительной работы сгибателей бедра во время фазы переноса во время спринта. Таким образом, эта благоприятная морфология может помочь достичь превосходных результатов в длинном спринте, возможно, за счет сохранения частоты шагов и обеспечения экономичного спринта у специализированных спринтеров на 400 м.
Напротив, отношение длины большеберцовой кости к длине бедренной кости коррелирует с оценкой ИААФ у спринтеров на 400 м, но не у спринтеров на 100 м. Таблица 2 Коэффициенты корреляции между переменными длины костей голени и результативностью в спринте (оценка Международной любительской легкоатлетической федерации) у специализированных спринтеров на 100 и 400 м более высокое отношение длины голени к длине бедренной кости коррелировало с лучшим результатом ИААФ у спринтеров на 400 м, но не у спринтеров на 100 м. Поддержание постоянной частоты шагов во время бега на 400 м необходимо для достижения превосходных результатов в длинном спринте [16, 23]. Морфологические факторы могут способствовать поддержанию частоты шагов, потенциально поддерживая экономичный спринт [5, 6]. Соотношение между длинами костей голени может быть полезным для уменьшения момента инерции ноги и положительной работы сгибателей бедра во время фазы переноса во время спринта. Таким образом, эта благоприятная морфология может помочь достичь превосходных результатов в длинном спринте, возможно, за счет сохранения частоты шагов и обеспечения экономичного спринта у специализированных спринтеров на 400 м.
Это исследование не выявило корреляции между отношением длины голени к длине бедра и оценкой ИААФ у спринтеров на 100 м. В целом, превосходные результаты в спринте на 100 м могут не требовать экономичного движения, потому что скорость спринта во время бега на 100 м существенно не снижается по сравнению со скоростью в спринте на 400 м [11, 24]. Кроме того, превосходные результаты в спринте на 100 м связаны с большей силой реакции опоры во время бега на 100 м [15, 40]. Увеличение силы опорной реакции при беге на 100 м может быть связано с большими размерами некоторых мышц ног из-за положительной связи между этими размерами мышц и результативностью в спринте на 100 м [1, 3, 7]. В частности, в предыдущих исследованиях было установлено, что более крупные мышцы бедра, включая четырехглавую мышцу бедра и подколенное сухожилие, коррелируют с лучшими результатами в спринте на 100 м у спринтеров [1, 3, 7]. При более высоком соотношении длины большеберцовой кости к длине бедра у спринтеров на 100 м эта морфология может моделировать меньшие мышцы бедра из-за необходимого укорочения длины бедра.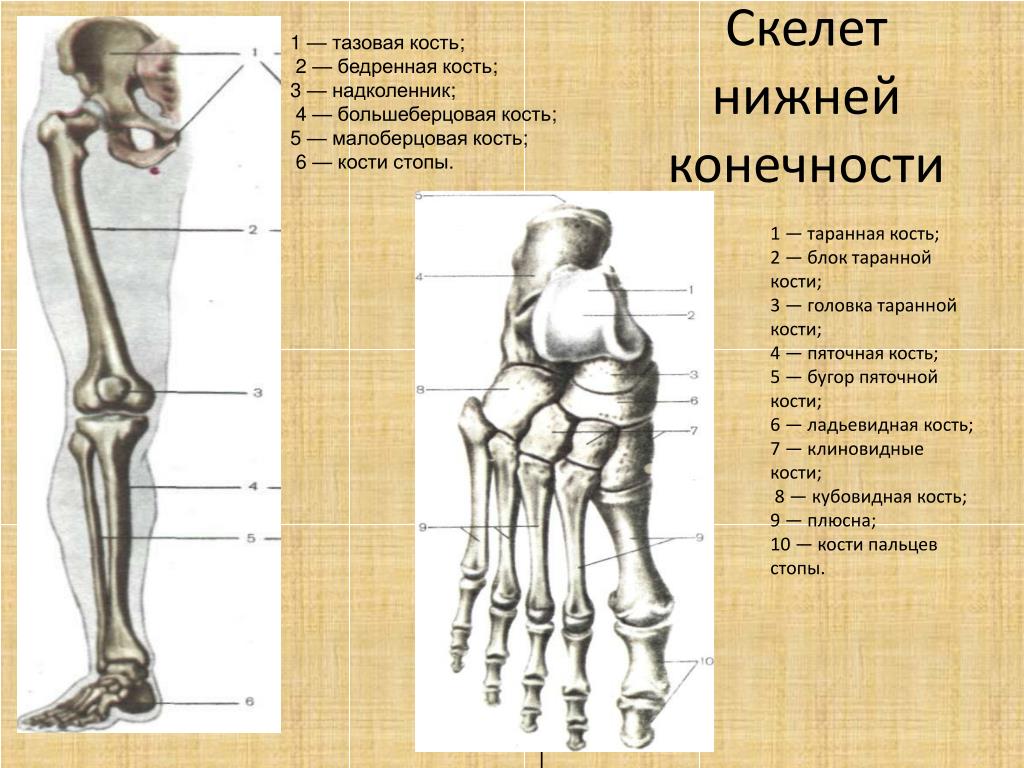 Следовательно, соотношение между длинами костей голени может не иметь отношения к результатам спринта на 100 м у специализированных спринтеров на 100 м.
Следовательно, соотношение между длинами костей голени может не иметь отношения к результатам спринта на 100 м у специализированных спринтеров на 100 м.
Ранее мы сообщали с помощью МРТ, что относительная длина костей голени (т. е. длина большеберцовой кости и общая длина бедренной и большеберцовой костей), нормализованные к росту, коррелирует с беговыми характеристиками у бегунов на выносливость [14]. Напротив, в настоящем исследовании абсолютная и относительная индивидуальная и общая длина костей голени не коррелировали с результатами спринта ни на 100 м, ни на 400 м. Хотя более длинная нога связана с увеличением длины шага при беге на короткие дистанции [9]., 10], длина шага может быть менее важным кинематическим фактором для достижения превосходных результатов в спринте, чем частота шагов при беге на 100 и 400 м [15,16,17]; в частности, увеличение длины шага не требуется для достижения превосходных результатов в спринте. Только одно исследование Morin et al. [15] сообщили, что относительная длина ноги (т.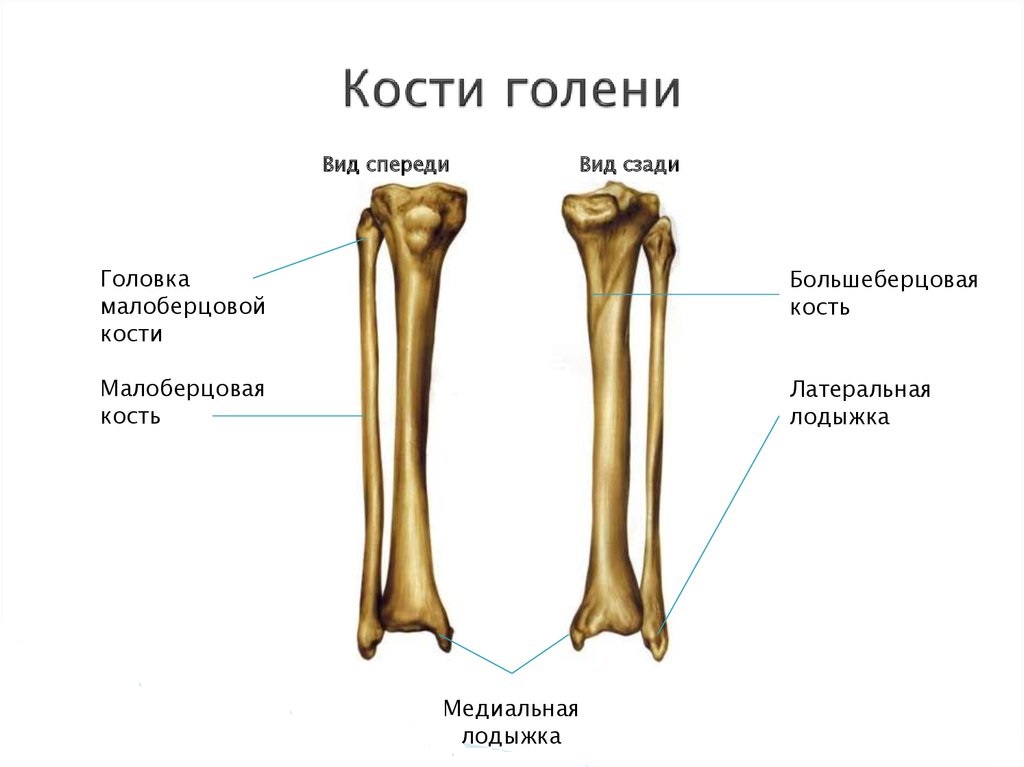 е. расстояние от большого вертела до земли), нормированная на рост тела, не коррелирует со скоростью бега на 100 м. Таким образом, настоящие результаты подтверждают этот результат, показывая отсутствие связи между длиной ноги и спринтерскими характеристиками у специализированных спринтеров на 100 м. Кроме того, настоящее исследование является первым, в котором установлено, что более длинная нога может не потребоваться для достижения превосходных результатов в длинном спринте у специализированных спринтеров на 400 м.
е. расстояние от большого вертела до земли), нормированная на рост тела, не коррелирует со скоростью бега на 100 м. Таким образом, настоящие результаты подтверждают этот результат, показывая отсутствие связи между длиной ноги и спринтерскими характеристиками у специализированных спринтеров на 100 м. Кроме того, настоящее исследование является первым, в котором установлено, что более длинная нога может не потребоваться для достижения превосходных результатов в длинном спринте у специализированных спринтеров на 400 м.
Это исследование показало, что все переменные длины костей не отличались между спринтерами на 100 и 400 м; таким образом, характеристики длины костей голени у этих двух групп одинаковы. В дополнительном анализе настоящего исследования абсолютная общая и индивидуальная длина бедра и голени у спринтеров (объединенная группа бегунов на 100 и 400 м) была выше, чем у бегунов на выносливость на 5000 м (например, 431,4 ± 13,9). по сравнению с 420,8 ± 20,2 мм для длины бедренной кости и 358,8 ± 13,8 по сравнению с 351,3 ± 18,2 мм для длины большеберцовой кости; P <0,05 для обоих), наблюдаемые в нашем предыдущем исследовании [14], что может быть связано с большей высотой тела у спринтеров, чем у бегунов на выносливость (172,8 ± 4,0 против 169,6 ± 5,6 см, P = 0,01). Напротив, относительная индивидуальная и общая длина бедренной и большеберцовой костей, а также отношение длины большеберцовой кости к длине бедренной кости не различались между спринтерами и бегунами на выносливость (25,0 ± 0,4 против 24,8 ± 0,7% обоих роста для относительной длины бедра, 20,8 ± 0,6 против 20,7 ± 0,6% роста относительной длины большеберцовой кости, 0,83 ± 0,02 против 0,84 ± 0,02; P > 0,05 для всех). Сходные результаты также наблюдались для переменных длины костей голени у спринтеров на 100 и 400 м и бегунов на выносливость (данные не показаны). Таким образом, характеристики длины костей голени относительно размера тела и соотношения между длинами костей голени могут быть сходными у спортсменов, соревнующихся на дистанциях от 100 м до 5000 м.
Напротив, относительная индивидуальная и общая длина бедренной и большеберцовой костей, а также отношение длины большеберцовой кости к длине бедренной кости не различались между спринтерами и бегунами на выносливость (25,0 ± 0,4 против 24,8 ± 0,7% обоих роста для относительной длины бедра, 20,8 ± 0,6 против 20,7 ± 0,6% роста относительной длины большеберцовой кости, 0,83 ± 0,02 против 0,84 ± 0,02; P > 0,05 для всех). Сходные результаты также наблюдались для переменных длины костей голени у спринтеров на 100 и 400 м и бегунов на выносливость (данные не показаны). Таким образом, характеристики длины костей голени относительно размера тела и соотношения между длинами костей голени могут быть сходными у спортсменов, соревнующихся на дистанциях от 100 м до 5000 м.
В наших предыдущих исследованиях было установлено, что более длинные кости передней части стопы и большее плечо момента разгибателя колена коррелируют с лучшими результатами в длинном спринте у спринтеров на 400 м [5, 6]. Насколько нам известно, никакие другие исследователи не сообщили о морфологических факторах, определяющих превосходные результаты в длинном спринте у спринтеров на 400 м. Эти благоприятные морфологические факторы превосходных результатов в спринте на 400 м, полученные в наших предыдущих исследованиях, также являются определяющими для достижения превосходных результатов в спринте на 100 м [2, 4]. Напротив, более высокое отношение длины голени к длине бедра может быть определяющим фактором только для спринтеров на 400 м. Таким образом, настоящее исследование является первым, в котором был обнаружен специфический морфологический фактор, способствующий превосходным результатам в длинном спринте у специализированных спринтеров на 400 м. Эта информация может быть полезна для выбора спринтерских видов и понимания индивидуальных особенностей спринтеров, особенно специализированных спринтеров на 400 м.
Насколько нам известно, никакие другие исследователи не сообщили о морфологических факторах, определяющих превосходные результаты в длинном спринте у спринтеров на 400 м. Эти благоприятные морфологические факторы превосходных результатов в спринте на 400 м, полученные в наших предыдущих исследованиях, также являются определяющими для достижения превосходных результатов в спринте на 100 м [2, 4]. Напротив, более высокое отношение длины голени к длине бедра может быть определяющим фактором только для спринтеров на 400 м. Таким образом, настоящее исследование является первым, в котором был обнаружен специфический морфологический фактор, способствующий превосходным результатам в длинном спринте у специализированных спринтеров на 400 м. Эта информация может быть полезна для выбора спринтерских видов и понимания индивидуальных особенностей спринтеров, особенно специализированных спринтеров на 400 м.
Ограничения
Мы предположили, что более высокое отношение длины большеберцовой кости к длине бедренной кости может помочь в достижении превосходных результатов в длинном спринте, потенциально за счет сохранения частоты шагов и выполнения экономичного спринта у специализированных спринтеров на 400 м. Однако мы не измеряли кинетические (например, силу реакции опоры) и кинематические (например, частоту шагов) данные во время бега на 400 м. Необходимы дальнейшие исследования для изучения взаимосвязи между переменными длины костей голени и кинетическими и кинематическими данными во время бега на 400 м.
Однако мы не измеряли кинетические (например, силу реакции опоры) и кинематические (например, частоту шагов) данные во время бега на 400 м. Необходимы дальнейшие исследования для изучения взаимосвязи между переменными длины костей голени и кинетическими и кинематическими данными во время бега на 400 м.
Наличие данных и материалов
Данные будут предоставлены соответствующим автором по запросу.
Сокращения
- ИААФ:
Международная любительская легкоатлетическая федерация
- МРТ:
Магнитно-резонансная томография
Ссылки
Ema R, Sakaguchi M, Kawakami Y. Большая мускулатура бедра и поясничной мышцы и ее связь с механикой бега у спринтеров. Медицинские спортивные упражнения. 2018;50(10):2085–91.
Артикул Google Scholar
 «>
«>Мияке Ю., Суга Т., Оцука М., Танака Т., Мисаки Дж., Кудо С., Нагано А., Исака Т. Момент плеча разгибателя колена связан с производительностью у мужчин-спринтеров. Eur J Appl Physiol. 2017;117(3):533–9.
Артикул Google Scholar
Сугисаки Н., Кобаяши К., Цучие Х., Канехиса Х. Связь между индивидуальными объемами мышц нижних конечностей и временем бега на 100 м у мужчин-спринтеров. Int J Sports Physiol Perform. 2018;13(2):214–9.
Артикул Google Scholar
Танака Т., Суга Т., Оцука М., Мисаки Дж., Мияке Й., Кудо С., Нагано А., Исака Т. Взаимосвязь между длиной костей переднего отдела стопы и результатами у мужчин-спринтеров. Scand J Med Sci Sports. 2017;27(12):1673–80.
Артикул КАС Google Scholar
Томита Д., Суга Т., Танака Т.
 , Уэно Х., Мияке Й., Оцука М., Нагано А., Исака Т. Пилотное исследование важности длины кости передней части стопы у мужчин, бегущих на 400 м на короткие дистанции: есть ли ключевой морфологический фактором для превосходных результатов в длинных спринтах? Примечания BMC Res. 2018;11:583.
, Уэно Х., Мияке Й., Оцука М., Нагано А., Исака Т. Пилотное исследование важности длины кости передней части стопы у мужчин, бегущих на 400 м на короткие дистанции: есть ли ключевой морфологический фактором для превосходных результатов в длинных спринтах? Примечания BMC Res. 2018;11:583.Артикул Google Scholar
Томита Д., Суга Т., Уэно Х., Мияке Й., Оцука М., Терада М., Нагано А., Исака Т. Взаимосвязь между плечом момента разгибателя колена и характеристиками длинного спринта у мужчин-спринтеров на 400 м. Транс Спорт Мед. 2018;1:172–9.
Артикул Google Scholar
Тоттори Н., Суга Т., Мияке Ю., Цутикане Р., Оцука М., Нагано А., Фудзита С., Исака Т. Мышечные мышцы сгибателей бедра и разгибателей колена связаны со спринтерскими характеристиками у тренированных спринтерских мальчиков подросткового возраста. Pediatr Exerc Sci.
 2018;30(1):115–23.
2018;30(1):115–23.Артикул Google Scholar
Weyand PG, Sternlight DB, Bellizzi MJ, Wright S. Более высокие максимальные скорости бега достигаются за счет большей силы на землю, а не более быстрых движений ног. J Appl Physiol. 2000;89(5):1991–99.
Артикул КАС Google Scholar
Cavanagh PR, Kram R. Длина шага в беге на длинные дистанции: скорость, размеры тела и дополнительные массовые эффекты. Медицинские спортивные упражнения. 1989;21(4):467–79.
Артикул КАС Google Scholar
Парузель-Дия М., Валажчик А., Искра Ю. Телосложение элитных спринтеров, длина и частота шагов. Конный физкультурный туризм. 2006;13(1):33–7.
Google Scholar
Макала К. Оптимизация производительности посредством кинематического анализа различных этапов бега на 100 метров.
 Новый Стад Спортсмен. 2007;22(2):7–16.
Новый Стад Спортсмен. 2007;22(2):7–16.Google Scholar
Laumets R, Viigipuu K, Mooses K, Mäestu J, Purge P, Pehme A, Kaasik P, Mooses M. Длина голени связана с экономичностью бега у кавказских бегунов высокого уровня. Джей Хам Кинет. 2017; 56: 229–39.
Артикул Google Scholar
Мусес М., Мусес К., Хайле Д.В., Дюруссель Дж., Каасик П., Питсиладис Ю.П. Диссоциация между экономичностью бега и эффективностью бега у элитных кенийских бегунов на длинные дистанции. J Sports Sci. 2015;33(2):136–44.
Артикул Google Scholar
Уэно Х., Суга Т., Такао К., Мияке Й., Терада М., Нагано А., Исака Т. Потенциальная взаимосвязь между длиной кости ноги и беговыми характеристиками у хорошо тренированных бегунов на выносливость. Джей Хам Кинет. 2019;70:165–72.

Артикул Google Scholar
Морин Дж.Б., Бурден М., Эдуард П., Пейро Н., Самозино П., Лакур Дж.Р. Механические детерминанты результативности бега на 100 м. Eur J Appl Physiol. 2012;112(11):3921–30.
Артикул Google Scholar
Хобара Х., Иноуэ К., Гоми К., Сакамото М., Мураока Т., Исо С., Каносуэ К. Непрерывное изменение пружинно-массовых характеристик во время спринта на 400 м. J Sci Med Sport. 2010;13(2):256–61.
Артикул Google Scholar
Манн Р., Герман Дж. Кинематический анализ результатов олимпийского спринта: мужчины на 200 метров. Int J Sport Biomech. 1985;1(2):151–62.
Артикул Google Scholar
Оцука М., Кавахара Т., Исака Т. Острая реакция хорошо тренированных спринтеров на забег на 100 м: более высокая скорость спринта достигается с увеличением частоты шагов по сравнению с скоростными тренировками.
 J Прочность Конд Рез. 2016;30(3):635–42.
J Прочность Конд Рез. 2016;30(3):635–42.Артикул Google Scholar
Hanon C, Lepretre PM, Bishop D, Thomas C. Потребление кислорода и метаболические реакции крови на бег на 400 м. Eur J Appl Physiol. 2010;109(2): 233–40.
Артикул КАС Google Scholar
Хилл Д.В. Вклад энергетической системы в бег на средние дистанции. J Sports Sci. 1999;17(6):477–83.
Артикул КАС Google Scholar
де Лева П. Корректировка параметров инерции участка Зациорского-Селуянова. Дж. Биомех. 1996;29(9):1223–30.
Артикул Google Scholar
Уэно Х., Суга Т., Такао К., Танака Т., Мисаки Дж., Мияке Й., Нагано А., Исака Т. Связь между длиной кости передней части стопы и результатами у бегунов на выносливость мужчин.
 Int J Sports Med. 2018;39(4):275–81.
Int J Sports Med. 2018;39(4):275–81.Артикул Google Scholar
Hanon C, Gajer B. Параметры скорости и шага бегунов на 400 метров мирового класса по сравнению с менее опытными бегунами. J Прочность Конд Рез. 2009;23(2):524–31.
Артикул Google Scholar
Славински Дж., Термоз Н., Рабита Г., Гилхем Г., Дорел С., Морин Дж. Б., Самозино П. Как анализ забега на 100 м улучшает наше понимание мужских и женских спринтерских результатов мирового класса. Scand J Med Sci Sports. 2017;27(1):45–54.
Артикул КАС Google Scholar
Скачать ссылки
Благодарности
Мы благодарны всем испытуемым, которые посвятили свое время и усилия для участия в этом исследовании.
Финансирование
Для настоящего исследования не было специальных грантов или финансирования.
Информация об авторе
Авторы и организации
Факультет спорта и здравоохранения, Университет Рицумейкан, 1-1-1 Нодзихигаси, Кусацу, Сига, 525-8577, Япония
Даичи Томита, Тадаси Суга, Масафуми Терада, Такахиро Танака, Юто Мияке, Хиромаса Уэно, Мицуо Оцука, Акинори Нагано и Тадао Исака
Высшая школа здоровья и спорта, Японский университет спортивных наук, Токио, Япония
Хиромаса Уэно
Научный сотрудник Японского общества содействия науке, Токио, Япония
Хиромаса Уэно
- Даичи Том ita
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Tadashi Suga
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Масафуми Терада
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Takahiro Tanaka
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Yuto Miyake
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Хиромаса Уэно
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Mitsuo Otsuka
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Akinori Nagano
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Тадао Исака
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
DT и TS задумали и разработали эксперимент; DT, TS, MT, TT, YM, HU и MO проводили эксперименты; данные анализа DT и TS; DT, TS, MT, TT, YM, HU MO, AN и TI интерпретировали результаты экспериментов; DT и TS написали рукопись; MT MO AN и TI отредактировали и переработали рукопись. Все авторы прочитали и одобрили рукопись.
Все авторы прочитали и одобрили рукопись.
Автор, ответственный за переписку
Переписка с Тадаши Суга.
Декларация этики
Утверждение этики и согласие на участие
Это исследование было одобрено Комитетом по этике Университета Рицумейкан (BKC-IRB-2011-009). Информированное письменное согласие было получено от всех участников.
Согласие на публикацию
Не применимо.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующую ссылку на оригинальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения.
