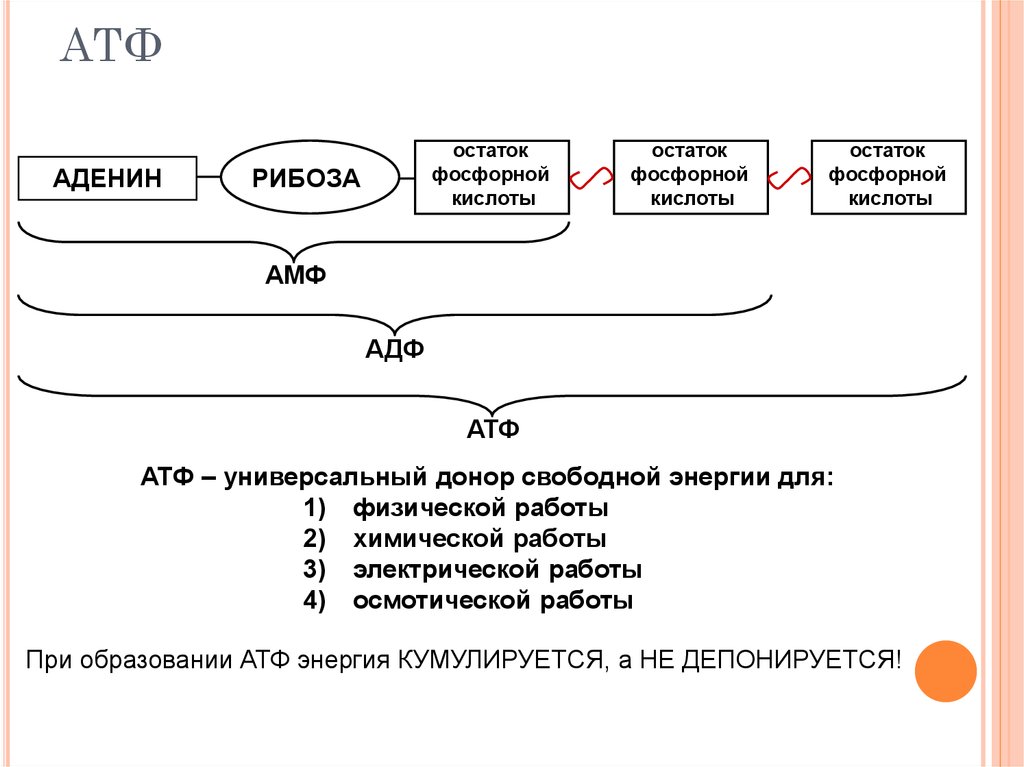
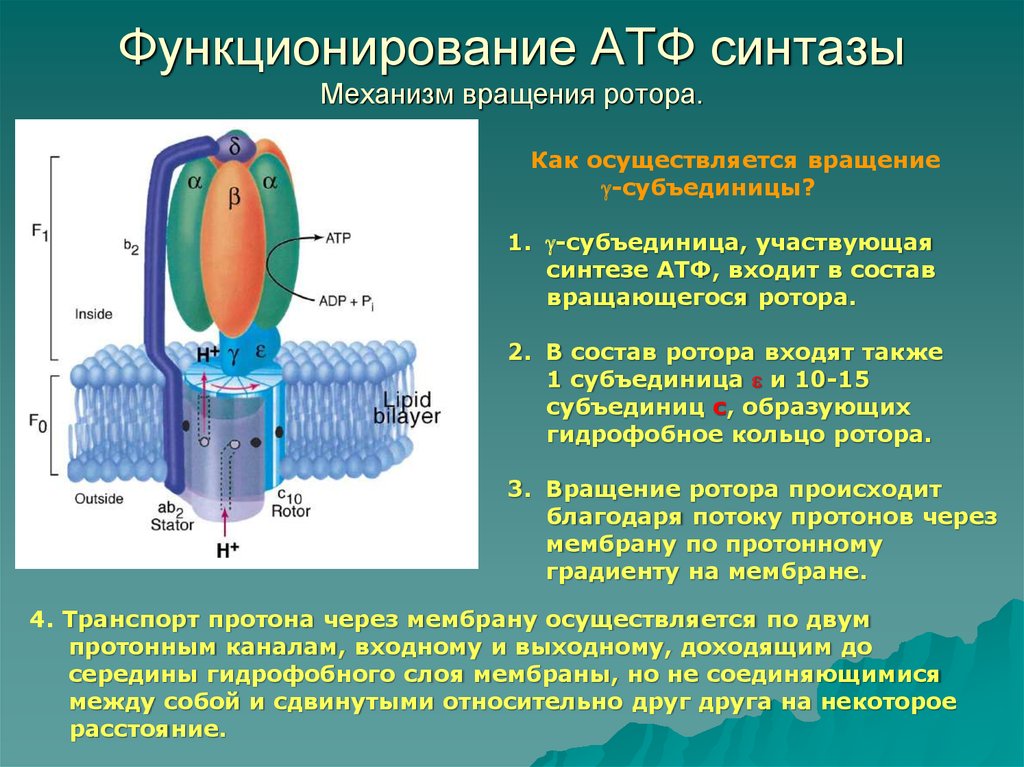
2. Строение атф-синтазы и синтез атф
АТФ-синтаза (Н+-АТФ-аза) — интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1 (рис. 6-15).
Рис.
6-15. Строение и механизм действия
АТФ-синтазы. А
— F0 и
F1 —
комплексы АТФ-синтазы, В состав F0входят
полипептидные цепи, которые образуют
канал, пронизывающий мембрану насквозь.
По этому каналу протоны возвращаются
в матрикс из межмембранного пространства;
белок F1 выступает
в матрикс с внутренней стороны мембраны
и содержит 9 субъединиц, 6 из которых
образуют 3 пары α и β («головка»),
прикрывающие стержневую часть, которая
состоит из 3 субъединиц γ, δ и ε. γ и ε
подвижны и образуют стержень, вращающийся
внутри неподвижной головки и связанный
с комплексом F0. В активных центрах,
образованных парами субъединиц α и β,
происходит связывание АДФ, неорганического
фосфата (Р Б — Каталитический цикл синтеза
АТФ включает 3 фазы, каждая из которых
проходит поочерёдно в 3 активных центрах:
1 — связывание АДФ и Н3РО4;
2 — образование фосфоангидридной связи
АТФ; 3 — освобождение конечного продукта.
При каждом переносе протонов через
канал F0 в
матрикс все 3 активных центра катализируют
очередную фазу цикла. Энергия
электрохимического потенциала расходуется
на поворот стержня, в результате которого
циклически изменяется конформация α-
и β-субъединиц и происходит синтез АТФ.
Б — Каталитический цикл синтеза
АТФ включает 3 фазы, каждая из которых
проходит поочерёдно в 3 активных центрах:
1 — связывание АДФ и Н3РО4;
2 — образование фосфоангидридной связи
АТФ; 3 — освобождение конечного продукта.
При каждом переносе протонов через
канал F0 в
матрикс все 3 активных центра катализируют
очередную фазу цикла. Энергия
электрохимического потенциала расходуется
на поворот стержня, в результате которого
циклически изменяется конформация α-
и β-субъединиц и происходит синтез АТФ.
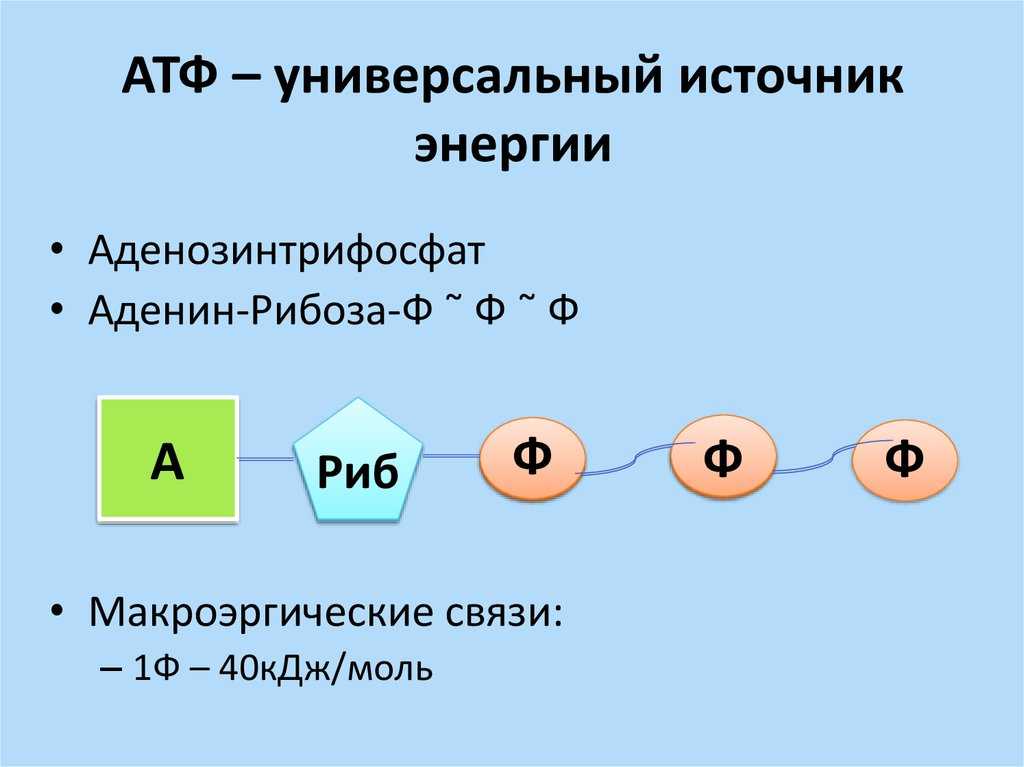
3.Коэффициент окислительного фосфорилирования
Окисление
молекулы NADH в ЦПЭ сопровождается
образованием 3 молекул АТФ; электроны
от FAD-зависимых дегидрогеназ поступают
в ЦПЭ на KoQ, минуя первый пункт сопряжения.
Поэтому образуются только 2 молекулы
АТФ. Отношение количества фосфорной
кислоты (Р), использованной на
фосфорилирование АДФ, к атому кислорода
(О), поглощённого в процессе дыхания,
называют коэффициентом окислительного
фосфорилирования и обозначают Р/О.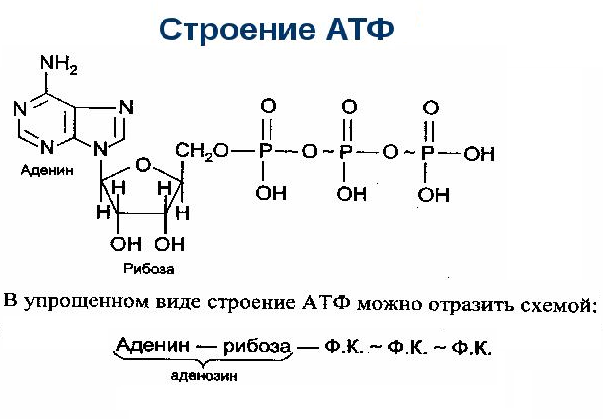
49.Регуляция цепи переноса электронов (дыхательный контроль). Разобщение тканевого дыхания и окислительного фосфорилирования. Терморегуляторная функция тканевого дыхания. Термогенная функция энергетического обмена в бурой жировой ткани.
4.Дыхательный контроль
Окисление
субстратов и фосфорилирование АДФ в
митохондриях прочно сопряжены. Скорость
использования АТФ регулирует скорость
потока электронов в ЦПЭ. Если АТФ не
используется и его концентрация в
клетках возрастает, то прекращается и
поток электронов к кислороду. С другой
стороны, расход АТФ и превращение его
в АДФ увеличивает окисление субстратов
и поглощение кислорода. Зависимость
интенсивности дыхания митохондрий от
концентрации АДФ называют дыхательным
контролем. Механизм дыхательного
контроля характеризуется высокой
точностью и имеет важное значение, так
как в результате его действия скорость
синтеза АТФ соответствует потребностям
клетки в энергии.
Общее содержание АТФ в организме 30-50 г, но каждая молекула АТФ в клетке «живёт» меньше минуты. В сутки у человека синтезируется 40-60 кг АТФ и столько же распадается. Увеличение концентрации АДФ немедленно приводит к ускорению дыхания и фосфорилирования.
Б. Транспорт АТФ и АДФ через мембраны митохондрий
В
большинстве эукариотических клеток
синтез основного количества АТФ
происходит внутри митохондрии, а основные
потребители АТФ расположены вне её. С
другой стороны, в матриксе митохондрий
должна поддерживаться достаточная
концентрация АДФ. Эти заряженные молекулы
не могут самостоятельно пройти через
липидный слой мембран. Внутренняя
мембрана непроницаема для заряженных
и гидрофильных веществ, но в ней содержится
определённое количество транспортёров,
избирательно переносящих подобные
молекулы из цитозоля в матрикс и из
матрикса в цитозоль.
В мембране есть белок АТФ/АДФ-антипортер, осуществляющий перенос этих метаболитов через мембрану (рис. 6-16). Молекула АДФ поступает в митоходриальный матрикс только при условии выхода молекулы АТФ из матрикса.
В. Разобщение дыхания и фосфорилирования
Некоторые
химические вещества (протонофоры) могут
переносить протоны или другие ионы
(ионофоры) из межмембранного пространства
через мембрану в матрикс, минуя протонные
каналы АТФ-синтазы.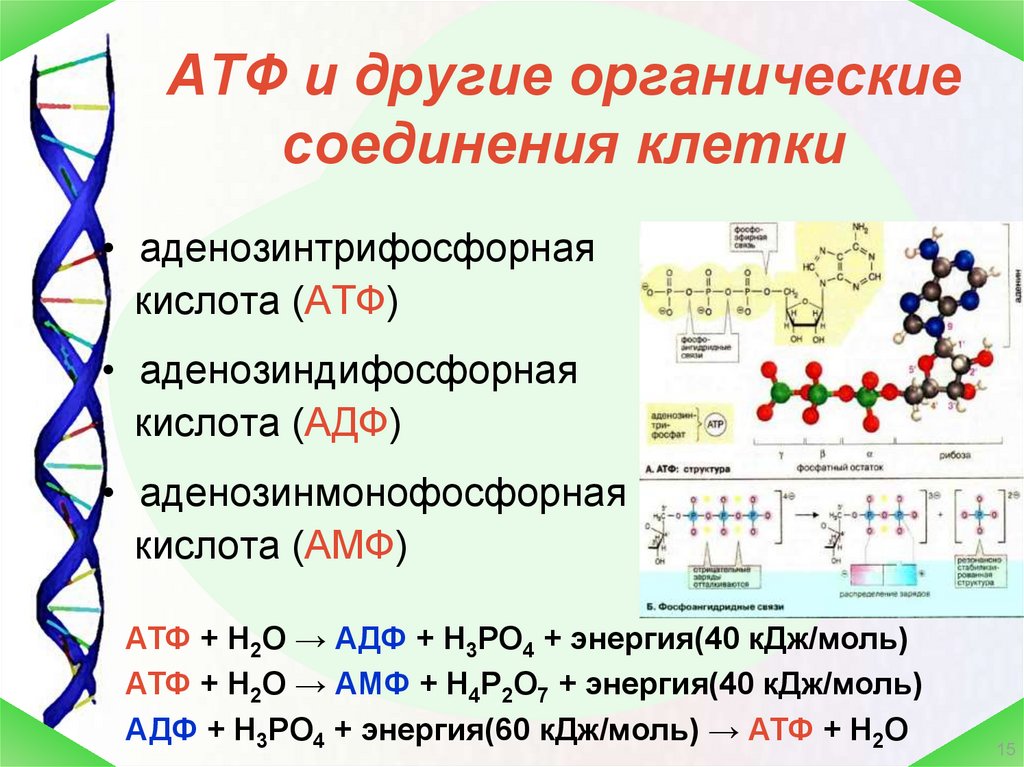 В результате этого
исчезает электрохимический потенциал
и прекращается синтез АТФ. Это явление
называют разобщением дыхания и
фосфорилирования. В результате разобщения
количество АТФ снижается, а АДФ
увеличивается. В этом случае скорость
окисления NADH и FADH 2возрастает,
возрастает и количество поглощённого
кислорода, но энергия выделяется в виде
теплоты, и коэффициент Р/О резко снижается.
Как правило, разобщители — липофильные
вещества, легко проходящие через липидный
слой мембраны. Одно из таких веществ —
2,4-динитрофенол, легко переходящий из
ионизированной формы в неионизированную,
присоединяя протон в межмембранном
пространстве и перенося его в матрикс.
В результате этого
исчезает электрохимический потенциал
и прекращается синтез АТФ. Это явление
называют разобщением дыхания и
фосфорилирования. В результате разобщения
количество АТФ снижается, а АДФ
увеличивается. В этом случае скорость
окисления NADH и FADH 2возрастает,
возрастает и количество поглощённого
кислорода, но энергия выделяется в виде
теплоты, и коэффициент Р/О резко снижается.
Как правило, разобщители — липофильные
вещества, легко проходящие через липидный
слой мембраны. Одно из таких веществ —
2,4-динитрофенол, легко переходящий из
ионизированной формы в неионизированную,
присоединяя протон в межмембранном
пространстве и перенося его в матрикс.
Примерами
разобщителей могут быть также некоторые
лекарства, например дикумарол —
антикоагулянт или метаболиты, которые
образуются в организме, билирубин —
продукт катаболизма тема, тироксин —
гормон щитовидной железы . Все эти
вещества проявляют разобщающее действие
только при их высокой концентрации.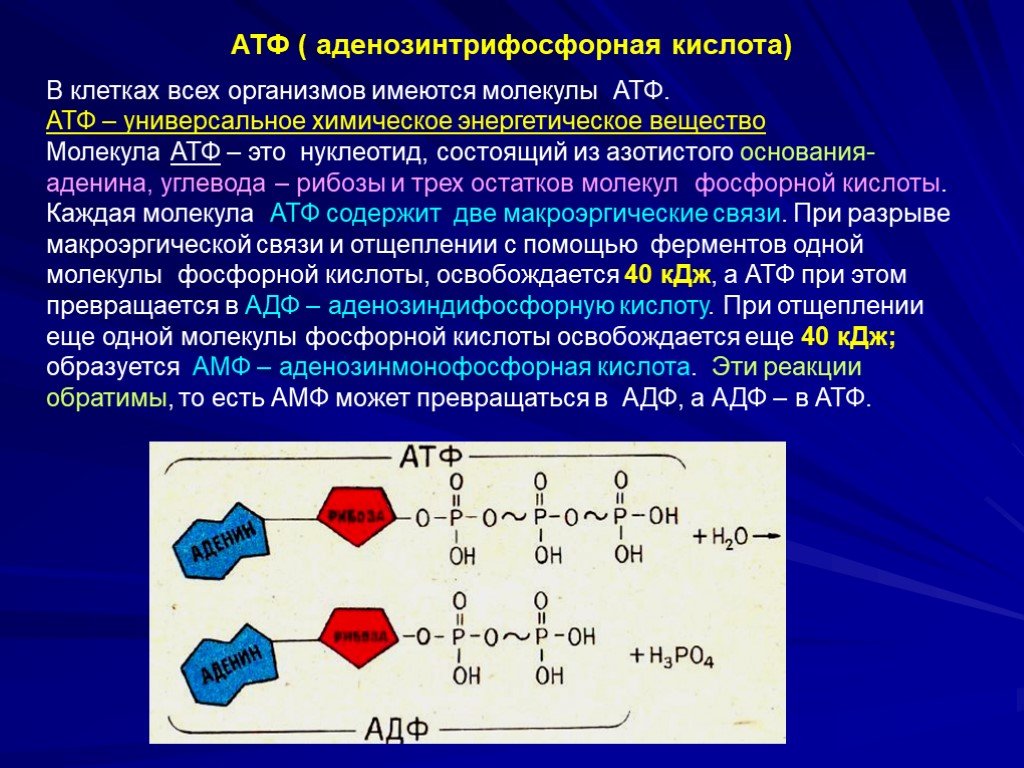
Г. Терморегуляторная функция ЦПЭ
На
синтез молекул АТФ расходуется примерно
40-45% всей энергии электронов, переносимых
по ЦПЭ, приблизительно 25% тратится на
работу по переносу веществ через
мембрану. Остальная часть энергии
рассеивается в виде теплоты и используется
теплокровными животными на поддержание
температуры тела. Кроме того, дополнительное
образование теплоты может происходить
при разобщении дыхания и фосфорилирования.
Разобщение окислительного фосфорилирования
может быть биологически полезным. Оно
позволяет генерировать тепло для
поддержания температуры тела у
новорождённых, у зимнеспящих животных
и у всех млекопитающих в процессе
адаптации к холоду. У новорождённых, а
также зимнеспящих животных существует
особая ткань, специализирующаяся на
теплопродукции посредством разобщения
дыхания и фосфорилирования — бурый жир.
Бурый жир содержит много митохондрий.
В мембране митохондрий имеется большой
избыток дыхательных ферментов по
сравнению с АТФ-синтазой.
На внешней стороне мембраны анион жирной кислоты присоединяет протон и в таком виде пересекает мембрану; на внутренней стороне мембраны диссоциирует, отдавая протон в матрикс и тем самым снижает протонный градиент. Образующийся анион возвращается на наружную сторону мембраны с помощью АТФ/ АДФ-антипортера.
 Образующиеся
свободные жирные кислоты служат не
только «топливом», но и важнейшим
регулятором разобщения дыхания и
фосфорилирования.
Образующиеся
свободные жирные кислоты служат не
только «топливом», но и важнейшим
регулятором разобщения дыхания и
фосфорилирования.АТФ-ЛОНГ (ATP-LONG). Таблетки пероральные Справочник лекарств. Томск
//= $single_post[«name_eng»] ?> //= $single_post[«name_rus»] ?>
Упаковка
Таблетки пероральныеФармакологическое действие
АТФ-ЛОНГ® является оригинальным лекарственным препаратом нового класса веществ – разнолигандных координационных соединений с макроэргическими фосфатами, молекула которого состоит из аденозин-5`-трифосфата (АТФ), аминокислоты гистидина и солей магния и калия. Благодаря оригинальной структуре молекула имеет характерную только для нее фармакологическое действие, которое не присуще любому из ее химических компонентов (АТФ, гистидин, К+, Mg++). АТФ-ЛОНГ® стимулирует энергетический обмен, активность ионтранспортных систем мембран клеток, нормализует концентрацию ионов калия и магния, показатели липидного состава мембран, активность мембранозависимых ферментов, улучшает антиоксидантную систему защиты миокарда, снижает концентрацию мочевой кислоты.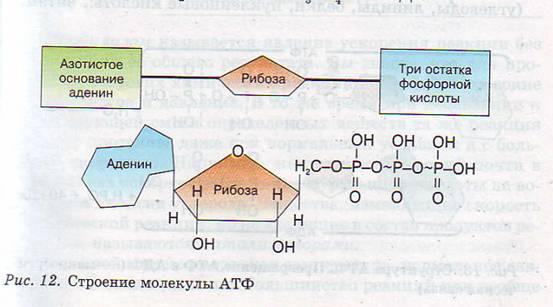 АТФ-ЛОНГ® обладает противоишемическим, мембраностабилизирующим, антиаритмическим действием благодаря нормализации метаболических процессов в миокарде при ишемии и гипоксии. АТФ-ЛОНГ® улучшает показатели центральной и периферической гемодинамики, коронарного кровообращения, повышает сократительную способность миокарда, функциональное состояние левого желудочка и сердечный выброс, который приводит к повышению показателей физической работоспособности. В условиях ишемии препарат уменьшает потребление миокардом кислорода, улучшает коронарное кровообращение, активирует функциональное состояние сердца, что приводит к уменьшению частоты приступов стенокардии и одышки во время физических нагрузок. Препарат восстанавливает нормальный синусовый ритм у больных пароксизмальной наджелудочковой и суправентрикулярной тахикардией, с мерцанием и трепетанием предсердий, а также уменьшает активность эктопических очагов (предсердные и желудочковые экстрасистолы). АТФ-ЛОНГ® нормализует концентрацию ионов калия и магния в тканях.
АТФ-ЛОНГ® обладает противоишемическим, мембраностабилизирующим, антиаритмическим действием благодаря нормализации метаболических процессов в миокарде при ишемии и гипоксии. АТФ-ЛОНГ® улучшает показатели центральной и периферической гемодинамики, коронарного кровообращения, повышает сократительную способность миокарда, функциональное состояние левого желудочка и сердечный выброс, который приводит к повышению показателей физической работоспособности. В условиях ишемии препарат уменьшает потребление миокардом кислорода, улучшает коронарное кровообращение, активирует функциональное состояние сердца, что приводит к уменьшению частоты приступов стенокардии и одышки во время физических нагрузок. Препарат восстанавливает нормальный синусовый ритм у больных пароксизмальной наджелудочковой и суправентрикулярной тахикардией, с мерцанием и трепетанием предсердий, а также уменьшает активность эктопических очагов (предсердные и желудочковые экстрасистолы). АТФ-ЛОНГ® нормализует концентрацию ионов калия и магния в тканях.
Показания к применению
Препарат назначается в комплексном лечении ишемической болезни сердца, нестабильной стенокардии, постинфарктного и миокардитического кардиосклероза, пароксизмальной наджелудочковой и суправентрикулярной тахикардии и в комплексной терапии других нарушений ритма; при вегетативных расстройствах, микрокардиодистрофии, синдроме хронической усталости, гиперурикемии различного происхождения.Форма выпуска
По 10 таблеток в контурной ячейковой упаковке; по 4 контурные упаковки в пачке. По 20 таблеток в контурной ячейковой упаковке; по 2 контурные упаковки в пачке.Фармакодинамика
АТФ-ЛОНГ® является оригинальным лекарственным препаратом нового класса веществ – разнолигандных координационных соединений с макроэргическими фосфатами, молекула которого состоит из аденозин-5`-трифосфата (АТФ), аминокислоты гистидина и солей магния и калия. Благодаря оригинальной структуре молекула имеет характерную только для нее фармакологическое действие, которое не присуще любому из ее химических компонентов (АТФ, гистидин, К+, Mg++). АТФ-ЛОНГ® стимулирует энергетический обмен, активность ионтранспортных систем мембран клеток, нормализует концентрацию ионов калия и магния, показатели липидного состава мембран, активность мембранозависимых ферментов, улучшает антиоксидантную систему защиты миокарда, снижает концентрацию мочевой кислоты. АТФ-ЛОНГ® обладает противоишемическим, мембраностабилизирующим, антиаритмическим действием благодаря нормализации метаболических процессов в миокарде при ишемии и гипоксии. АТФ-ЛОНГ® улучшает показатели центральной и периферической гемодинамики, коронарного кровообращения, повышает сократительную способность миокарда, функциональное состояние левого желудочка и сердечный выброс, который приводит к повышению показателей физической работоспособности. В условиях ишемии препарат уменьшает потребление миокардом кислорода, улучшает коронарное кровообращение, активирует функциональное состояние сердца, что приводит к уменьшению частоты приступов стенокардии и одышки во время физических нагрузок.
АТФ-ЛОНГ® стимулирует энергетический обмен, активность ионтранспортных систем мембран клеток, нормализует концентрацию ионов калия и магния, показатели липидного состава мембран, активность мембранозависимых ферментов, улучшает антиоксидантную систему защиты миокарда, снижает концентрацию мочевой кислоты. АТФ-ЛОНГ® обладает противоишемическим, мембраностабилизирующим, антиаритмическим действием благодаря нормализации метаболических процессов в миокарде при ишемии и гипоксии. АТФ-ЛОНГ® улучшает показатели центральной и периферической гемодинамики, коронарного кровообращения, повышает сократительную способность миокарда, функциональное состояние левого желудочка и сердечный выброс, который приводит к повышению показателей физической работоспособности. В условиях ишемии препарат уменьшает потребление миокардом кислорода, улучшает коронарное кровообращение, активирует функциональное состояние сердца, что приводит к уменьшению частоты приступов стенокардии и одышки во время физических нагрузок.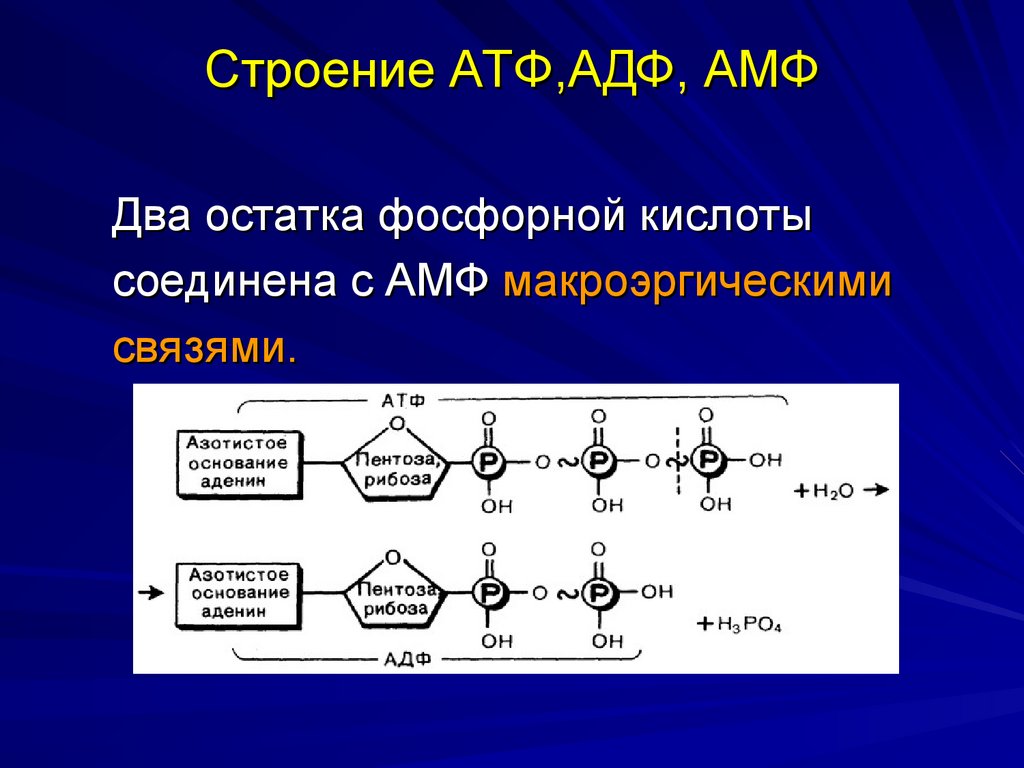 Препарат восстанавливает нормальный синусовый ритм у больных пароксизмальной наджелудочковой и суправентрикулярной тахикардией, с мерцанием и трепетанием предсердий, а также уменьшает активность эктопических очагов (предсердные и желудочковые экстрасистолы). АТФ-ЛОНГ® нормализует концентрацию ионов калия и магния в тканях.
Препарат восстанавливает нормальный синусовый ритм у больных пароксизмальной наджелудочковой и суправентрикулярной тахикардией, с мерцанием и трепетанием предсердий, а также уменьшает активность эктопических очагов (предсердные и желудочковые экстрасистолы). АТФ-ЛОНГ® нормализует концентрацию ионов калия и магния в тканях.Фармакокинетика
Фармакокинетика не изучалась.Противопоказания к применению
Острый инфаркт миокарда, тяжелые формы бронхиальной астмы, гиперкалиемия, гипермагниемия, повышенная чувствительность к компонентам препарата. AV блокады II-III степени, геморрагический инсульт, беременность, период кормления грудью.Побочные действия
Не установлено. Возможно появление тошноты, неприятных ощущений в эпигастрии, при продолжительном и неконтролируемом приеме — гиперкалиемия или гипермагниемия. Аллергические реакции.Способ применения и дозировка
Таблетки АТФ-ЛОНГ® принимают сублингвально (под язык) и держат до полного рассасывания. Разовая доза — 10-40 мг 3-4 раза в день, независимо от приема пищи.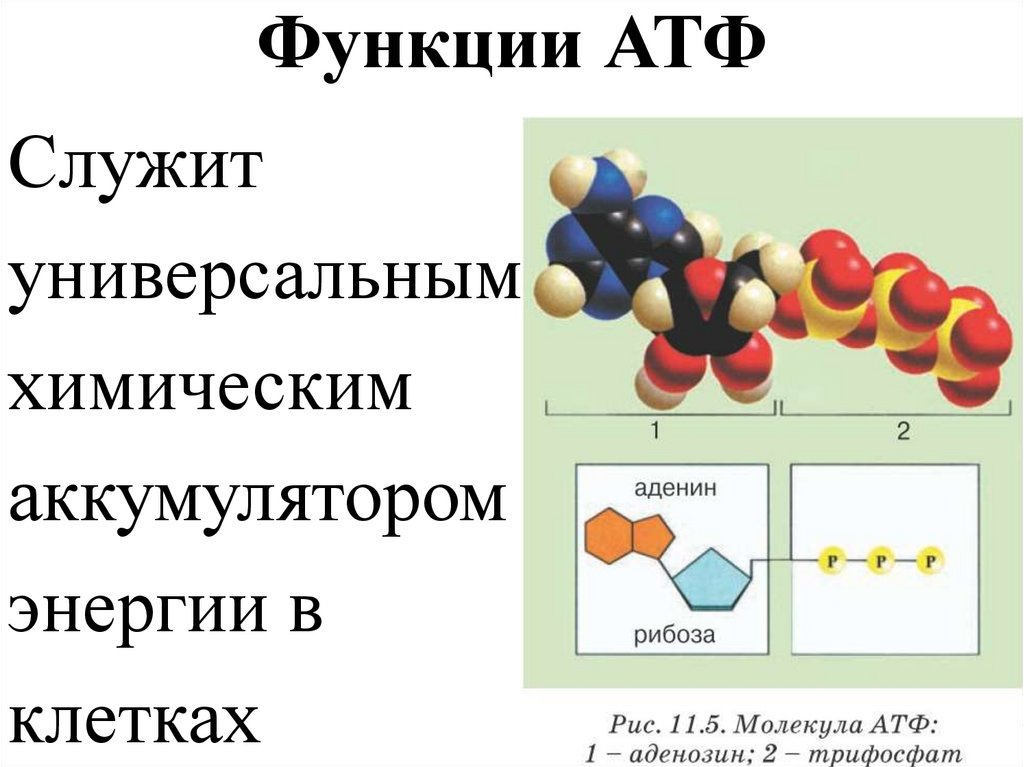 Срок лечения определяет врач, в среднем он составляет 20-30 дней. При необходимости курс повторяют через 10-15 дней. При острых сердечных состояниях и пароксизмальной мерцательной аритмии разовую дозу 10-40 мг принимают через каждые 5-10 минут до исчезновения негативных симптомов. Максимальная суточная доза составляет 400-600 мг. Опыта применения препарата у детей нет.
Срок лечения определяет врач, в среднем он составляет 20-30 дней. При необходимости курс повторяют через 10-15 дней. При острых сердечных состояниях и пароксизмальной мерцательной аритмии разовую дозу 10-40 мг принимают через каждые 5-10 минут до исчезновения негативных симптомов. Максимальная суточная доза составляет 400-600 мг. Опыта применения препарата у детей нет.Передозировка
Возможно развитие брадикардии, при которой необходимо введение атропина сульфата. Также возможно возникновение AV блокады и артериальной гипотензии.Взаимодействие с другими препаратами
АТФ-ЛОНГ® нельзя применять одновременно с сердечными гликозидами в связи повышением риска возникновения атриовентрикулярной блокады. При одновременном применении с калийсберегающими диуретиками и ингибиторами АПФ повышается риск развития гиперкалиемии, с магнеротом — гипермагниемии. Дипиридамол усиливает терапевтический эффект АТФ-ЛОНГ®а, а ксантинола никотинат – уменьшает.Особые указания при приеме
При сочетании атриовентрикулярной блокады с другими нарушениями ритма препарат не назначают. При продолжительном применении АТФ-ЛОНГ®а необходимо контролировать уровень калия и магния в крови.
При продолжительном применении АТФ-ЛОНГ®а необходимо контролировать уровень калия и магния в крови.Условия хранения
Хранить в недоступном для детей, сухом, защищенном от света месте при температуре от 3 до 5 °С.Срок годности
12 мес.Физиология, Аденозинтрифосфат — StatPearls
Введение
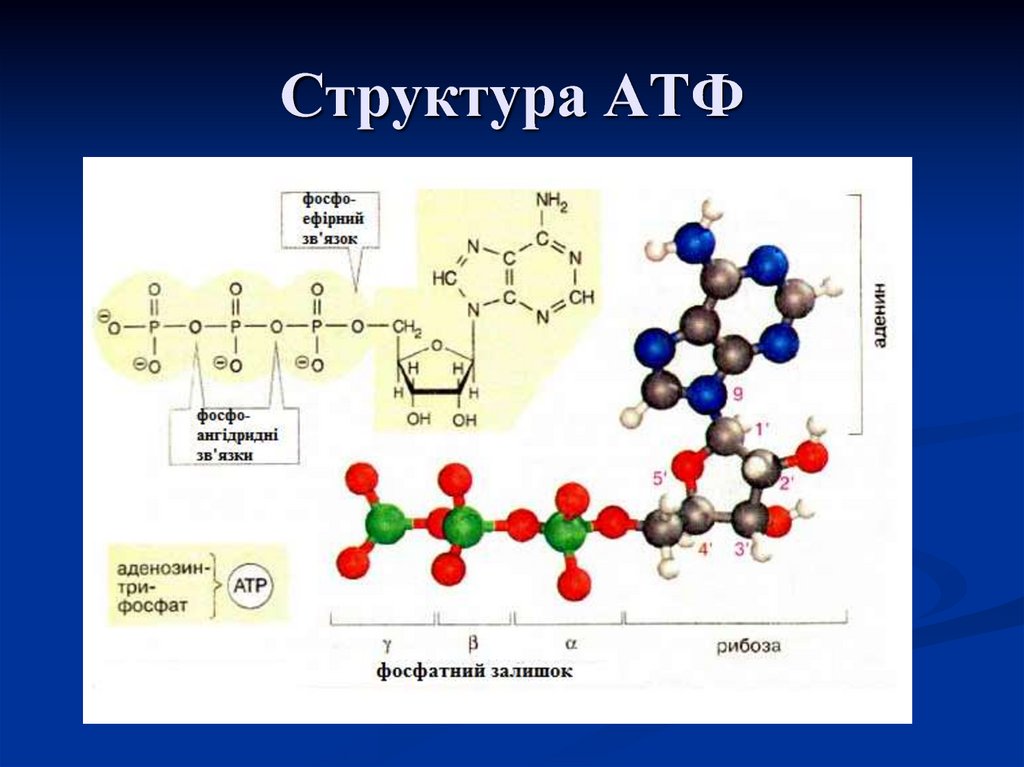
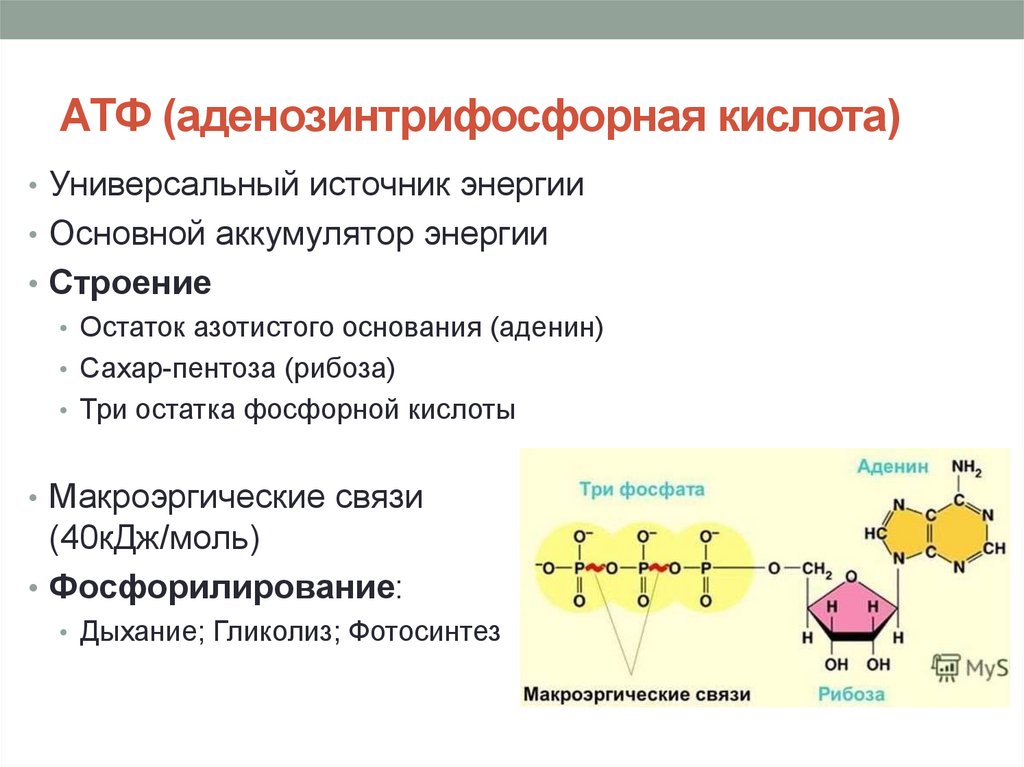
Тело представляет собой сложный организм, и поэтому ему требуется энергия для поддержания нормального функционирования. Аденозинтрифосфат (АТФ) является источником энергии для использования и хранения на клеточном уровне. Структура АТФ представляет собой нуклеозидтрифосфат, состоящий из азотистого основания (аденина), сахара рибозы и трех последовательно связанных фосфатных групп. АТФ обычно называют «энергетической валютой» клетки, поскольку он обеспечивает легко высвобождаемую энергию в связи между второй и третьей фосфатными группами. Помимо обеспечения энергией, расщепление АТФ посредством гидролиза выполняет широкий спектр клеточных функций, включая передачу сигналов и синтез ДНК/РНК.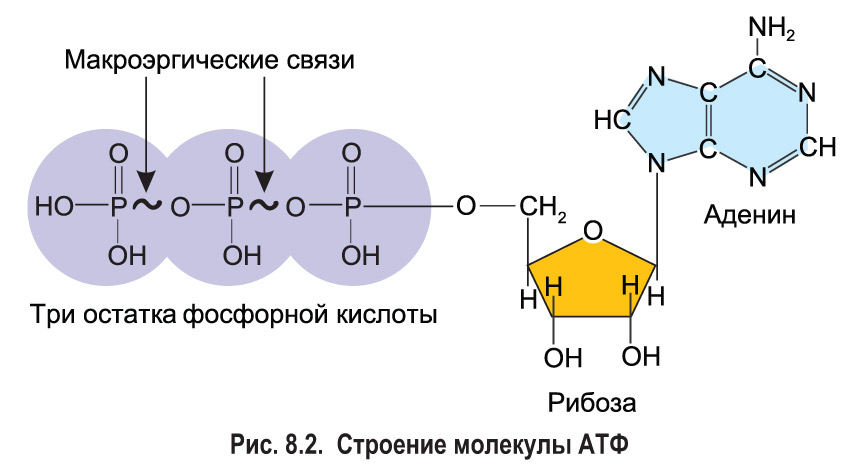
Большая часть синтеза АТФ происходит при клеточном дыхании в митохондриальном матриксе: образуется примерно тридцать две молекулы АТФ на молекулу окисляемой глюкозы. АТФ потребляется для получения энергии в процессах, включая перенос ионов, сокращение мышц, распространение нервных импульсов, фосфорилирование субстрата и химический синтез. Эти процессы, как и другие, создают высокий спрос на АТФ. В результате клетки в организме человека зависят от гидролиза от 100 до 150 моль АТФ в день для обеспечения правильного функционирования. В следующих разделах будет проведена дальнейшая оценка роли АТФ как ключевой молекулы в ежедневном функционировании клетки.
Сотовый уровень
АТФ является отличным накопителем энергии для использования в качестве «валюты» благодаря фосфатным группам, которые соединяются посредством фосфодиэфирных связей. Эти связи имеют высокую энергию из-за связанных с ними электроотрицательных зарядов, создающих силу отталкивания между фосфатными группами. Значительное количество энергии остается запасенным в фосфатно-фосфатных связях. В ходе метаболических процессов АТФ гидролизуется до АДФ или далее до АМФ и свободных неорганических фосфатных групп. Процесс гидролиза АТФ до АДФ энергетически выгоден, давая свободную энергию Гиббса -7,3 кал/моль.[1] АТФ должен постоянно пополняться, чтобы питать постоянно работающую клетку. Обычная внутриклеточная концентрация АТФ составляет от 1 до 10 мкМ.[2] Существует множество механизмов обратной связи, обеспечивающих поддержание постоянного уровня АТФ в клетке. Усиление или ингибирование АТФ-синтазы является обычным регуляторным механизмом. Например, АТФ ингибирует фосфофруктокиназу-1 (PFK1) и пируваткиназу, два ключевых фермента гликолиза, эффективно действуя как петля отрицательной обратной связи для ингибирования распада глюкозы при наличии достаточного количества клеточного АТФ.
Эти связи имеют высокую энергию из-за связанных с ними электроотрицательных зарядов, создающих силу отталкивания между фосфатными группами. Значительное количество энергии остается запасенным в фосфатно-фосфатных связях. В ходе метаболических процессов АТФ гидролизуется до АДФ или далее до АМФ и свободных неорганических фосфатных групп. Процесс гидролиза АТФ до АДФ энергетически выгоден, давая свободную энергию Гиббса -7,3 кал/моль.[1] АТФ должен постоянно пополняться, чтобы питать постоянно работающую клетку. Обычная внутриклеточная концентрация АТФ составляет от 1 до 10 мкМ.[2] Существует множество механизмов обратной связи, обеспечивающих поддержание постоянного уровня АТФ в клетке. Усиление или ингибирование АТФ-синтазы является обычным регуляторным механизмом. Например, АТФ ингибирует фосфофруктокиназу-1 (PFK1) и пируваткиназу, два ключевых фермента гликолиза, эффективно действуя как петля отрицательной обратной связи для ингибирования распада глюкозы при наличии достаточного количества клеточного АТФ.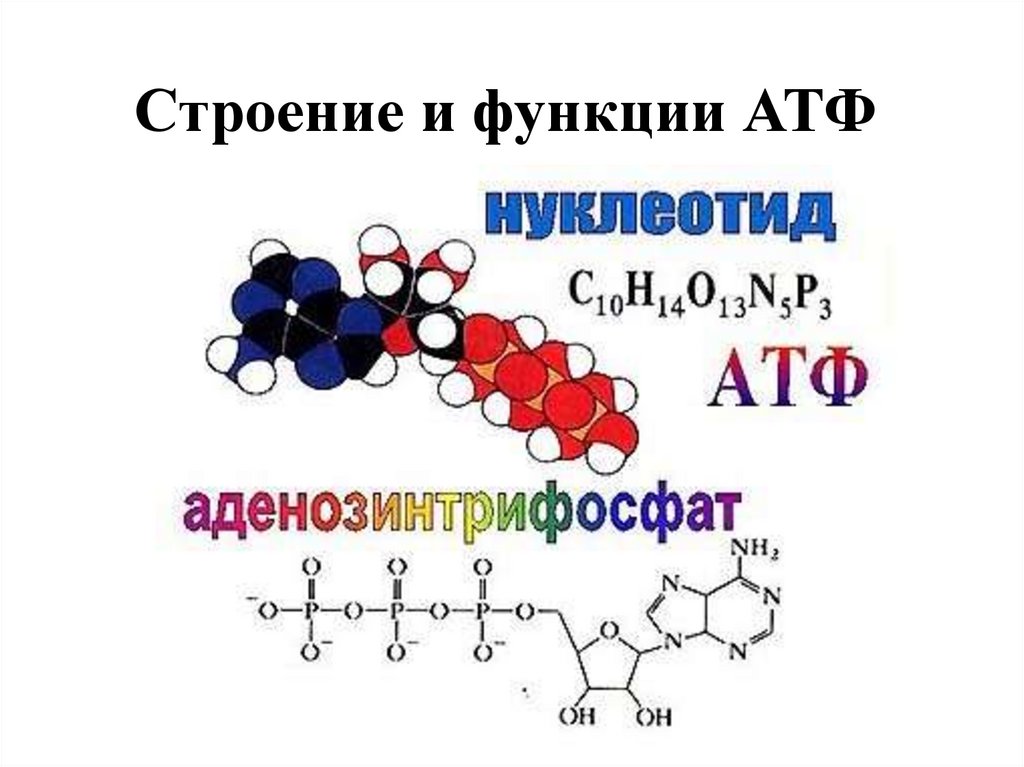
И наоборот, АДФ и АМФ могут активировать PFK1 и пируваткиназу, способствуя синтезу АТФ в периоды высокой потребности в энергии. Другие системы регулируют АТФ, например, в регуляторных механизмах, участвующих в регуляции синтеза АТФ в сердце. Новые эксперименты показали, что десятисекундные всплески, называемые митохондриальными вспышками, могут нарушить выработку АТФ в сердце. Во время этих митохондриальных вспышек митохондрии выделяют активные формы кислорода и эффективно приостанавливают синтез АТФ. Ингибирование продукции АТФ происходит во время митохондриальных вспышек. Во время низкой потребности в энергии, когда клетки сердечной мышцы получали достаточно строительных блоков, необходимых для производства АТФ, митохондриальные вспышки наблюдались чаще. С другой стороны, когда потребность в энергии высока во время быстрого сердечного сокращения, митохондриальные вспышки происходили реже. Эти результаты свидетельствуют о том, что в периоды, когда необходимы значительные количества АТФ, митохондриальные вспышки происходят реже, чтобы обеспечить продолжение производства АТФ.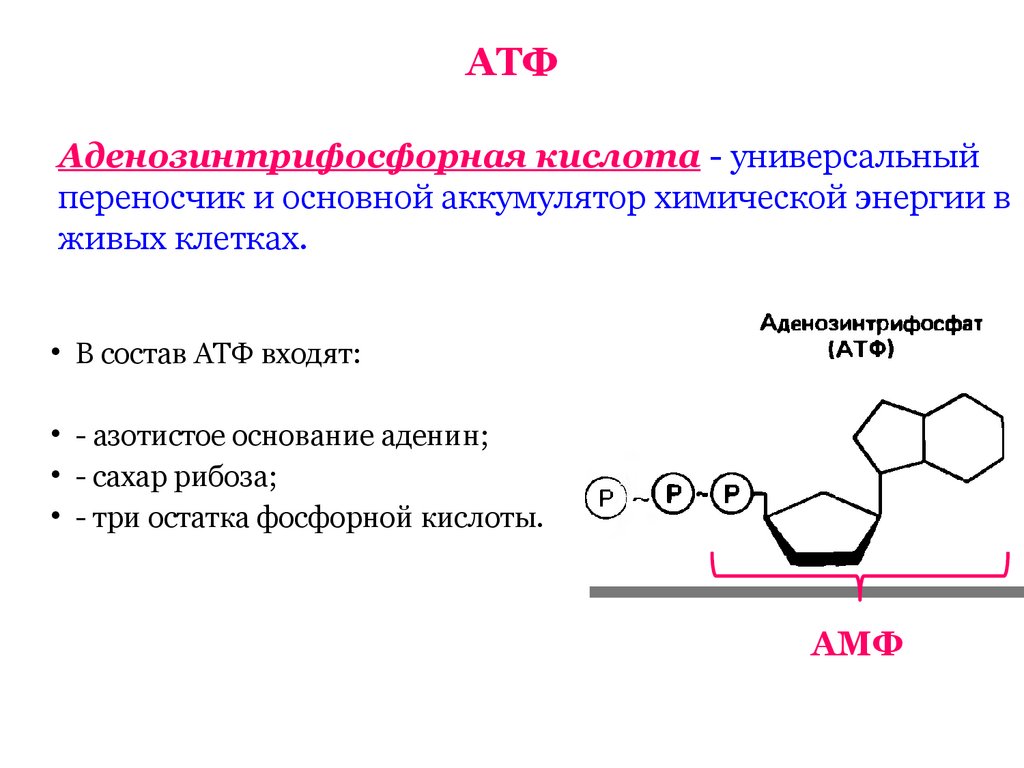 И наоборот, в периоды низкой выработки энергии митохондриальные вспышки происходили более регулярно и ингибировали выработку АТФ.[3]
И наоборот, в периоды низкой выработки энергии митохондриальные вспышки происходили более регулярно и ингибировали выработку АТФ.[3]
Функция
Гидролиз АТФ обеспечивает энергию, необходимую для многих важных процессов в организмах и клетках. К ним относятся внутриклеточная передача сигналов, синтез ДНК и РНК, пуринергическая передача сигналов, синаптическая передача сигналов, активный транспорт и сокращение мышц. Эти темы не являются исчерпывающим списком, но включают в себя некоторые жизненно важные роли, которые выполняет ATP.
АТФ во внутриклеточной передаче сигналов
Передача сигнала в значительной степени зависит от АТФ. АТФ может служить субстратом для киназ, самого многочисленного АТФ-связывающего белка. Когда киназа фосфорилирует белок, может активироваться сигнальный каскад, что приводит к модуляции различных внутриклеточных сигнальных путей.[4] Активность киназы жизненно важна для клетки и, следовательно, должна жестко регулироваться. Присутствие иона магния помогает регулировать активность киназы.[5] Регуляция осуществляется через ионы магния, находящиеся в клетке в виде комплекса с АТФ, связанного на фосфатно-кислородных центрах. В дополнение к киназной активности АТФ может функционировать как повсеместный триггер высвобождения внутриклеточного мессенджера.[6] Эти мессенджеры включают гормоны, различные ферменты, липидные медиаторы, нейротрансмиттеры, оксид азота, факторы роста и активные формы кислорода.[6] Пример использования АТФ во внутриклеточной передаче сигналов можно наблюдать в АТФ, выступающем в качестве субстрата для аденилатциклазы. Этот процесс в основном происходит в сигнальных путях рецепторов, связанных с G-белком. При связывании с аденилатциклазой АТФ превращается в циклический АМФ, который способствует высвобождению кальция из внутриклеточных запасов.[7] У цАМФ есть и другие роли, в том числе вторичные мессенджеры в сигнальных каскадах гормонов, активация протеинкиназ и регуляция функции ионных каналов.
Присутствие иона магния помогает регулировать активность киназы.[5] Регуляция осуществляется через ионы магния, находящиеся в клетке в виде комплекса с АТФ, связанного на фосфатно-кислородных центрах. В дополнение к киназной активности АТФ может функционировать как повсеместный триггер высвобождения внутриклеточного мессенджера.[6] Эти мессенджеры включают гормоны, различные ферменты, липидные медиаторы, нейротрансмиттеры, оксид азота, факторы роста и активные формы кислорода.[6] Пример использования АТФ во внутриклеточной передаче сигналов можно наблюдать в АТФ, выступающем в качестве субстрата для аденилатциклазы. Этот процесс в основном происходит в сигнальных путях рецепторов, связанных с G-белком. При связывании с аденилатциклазой АТФ превращается в циклический АМФ, который способствует высвобождению кальция из внутриклеточных запасов.[7] У цАМФ есть и другие роли, в том числе вторичные мессенджеры в сигнальных каскадах гормонов, активация протеинкиназ и регуляция функции ионных каналов.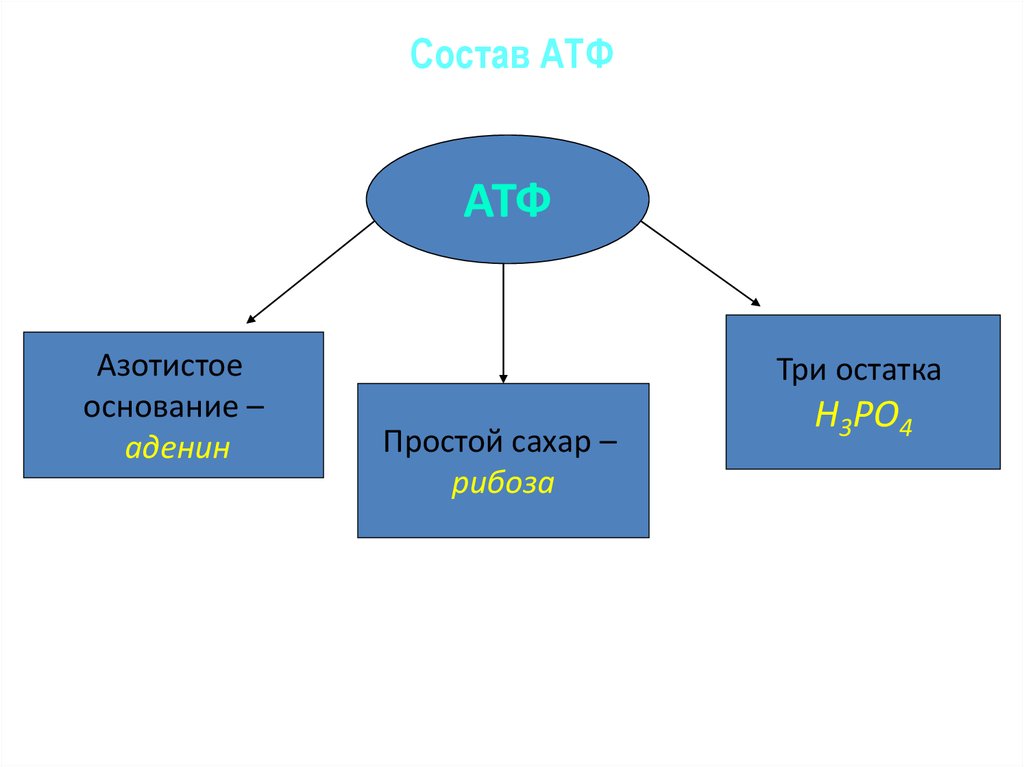
Синтез ДНК/РНК
Для синтеза ДНК и РНК требуется АТФ. АТФ является одним из четырех нуклеотид-трифосфатных мономеров, необходимых при синтезе РНК. Синтез ДНК использует аналогичный механизм, за исключением того, что при синтезе ДНК АТФ сначала трансформируется путем удаления атома кислорода из сахара с образованием дезоксирибонуклеотида, дАТФ.
Пуринергическая сигнализация
Пуринергическая передача сигналов представляет собой форму внеклеточной паракринной передачи сигналов, которая опосредована пуриновыми нуклеотидами, включая АТФ. Этот процесс обычно влечет за собой активацию пуринергических рецепторов на клетках в непосредственной близости, тем самым передавая сигналы для регулирования внутриклеточных процессов. АТФ высвобождается из везикулярных запасов и регулируется IP3 и другими распространенными экзоцитотическими регуляторными механизмами. АТФ хранится и высвобождается вместе с нейротрансмиттерами, что еще раз подтверждает представление о том, что АТФ является необходимым медиатором пуринергической нейротрансмиссии как в симпатических, так и в парасимпатических нервах. АТФ может индуцировать несколько пуринергических реакций, включая контроль вегетативных функций, взаимодействие нервной глии, боль и контроль тонуса сосудов.[9]][10][11][12]
АТФ может индуцировать несколько пуринергических реакций, включая контроль вегетативных функций, взаимодействие нервной глии, боль и контроль тонуса сосудов.[9]][10][11][12]
Нейротрансмиссия
Мозг является самым большим потребителем АТФ в организме, потребляя примерно двадцать пять процентов всей доступной энергии.[13] Большое количество энергии тратится на поддержание концентрации ионов для правильной передачи сигналов нейронами и синаптической передачи.[14] Синаптическая передача является энергоемким процессом. На пресинаптических окончаниях АТФ требуется для установления ионного градиента, который перемещает нейротрансмиттеры в везикулы, и для подготовки везикул к высвобождению посредством экзоцитоза. [14] Нейрональная передача сигналов зависит от потенциала действия, достигающего пресинаптических окончаний, сигнализирующего о высвобождении загруженных везикул. Этот процесс зависит от того, что АТФ восстанавливает концентрацию ионов в аксоне после каждого потенциала действия, позволяя возникать другому сигналу.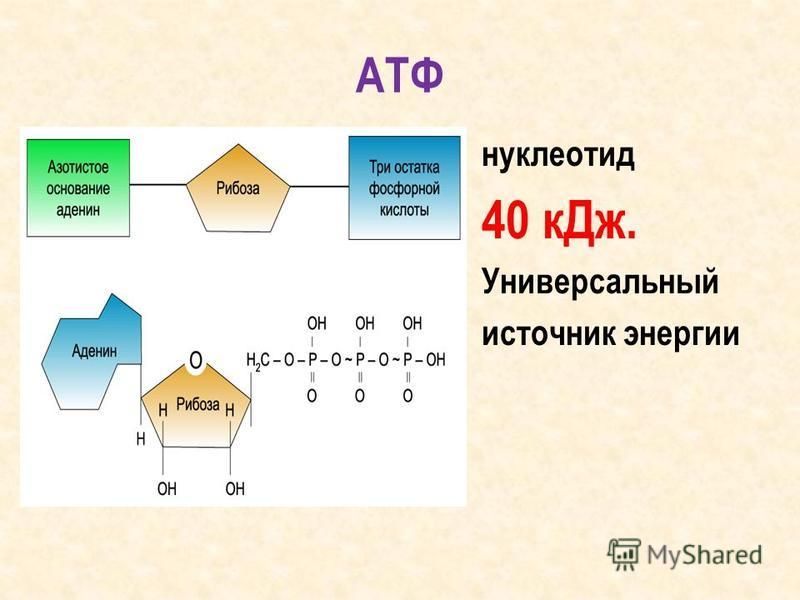 Активный транспорт отвечает за сброс концентраций ионов натрия и калия до исходных значений после возникновения потенциала действия через Na/K-АТФазу. Во время этого процесса одна молекула АТФ гидролизуется, три иона натрия транспортируются из клетки и два иона калия транспортируются обратно в клетку, оба из которых движутся против градиента их концентрации.
Активный транспорт отвечает за сброс концентраций ионов натрия и калия до исходных значений после возникновения потенциала действия через Na/K-АТФазу. Во время этого процесса одна молекула АТФ гидролизуется, три иона натрия транспортируются из клетки и два иона калия транспортируются обратно в клетку, оба из которых движутся против градиента их концентрации.
Потенциалы действия, проходящие по аксону, инициируют везикулярное высвобождение при достижении пресинаптического окончания. После установления ионного градиента потенциалы действия затем распространяются вниз по аксону посредством деполяризации аксона, посылая сигнал к терминалу. Приблизительно один миллиард ионов натрия необходим для распространения одного потенциала действия. Нейронам потребуется гидролизовать около миллиарда молекул АТФ, чтобы восстановить концентрацию ионов натрия/калия после каждой клеточной деполяризации.[13]Возбуждающие синапсы в значительной степени преобладают в сером веществе мозга. Везикулы, содержащие глутамат, высвобождаются в синаптическую щель, чтобы активировать постсинаптические возбуждающие глутаминергические рецепторы. Для загрузки этих молекул требуется большое количество АТФ из-за почти четырех тысяч молекул глутамата, хранящихся в одном пузырьке. Необходимы значительные запасы энергии, чтобы инициировать высвобождение везикул, управлять глутаматергическими постсинаптическими процессами и перерабатывать везикулы, а также оставшийся глутамат. Следовательно, из-за большого количества энергии, необходимой для упаковки глутамата, митохондрии близки к глутаматергическим пузырькам.
Для загрузки этих молекул требуется большое количество АТФ из-за почти четырех тысяч молекул глутамата, хранящихся в одном пузырьке. Необходимы значительные запасы энергии, чтобы инициировать высвобождение везикул, управлять глутаматергическими постсинаптическими процессами и перерабатывать везикулы, а также оставшийся глутамат. Следовательно, из-за большого количества энергии, необходимой для упаковки глутамата, митохондрии близки к глутаматергическим пузырькам.
АТФ в сокращении мышц
Сокращение мышц является необходимой функцией повседневной жизни и не может происходить без АТФ. Есть три основные роли, которые АТФ выполняет в действии мышечного сокращения. Во-первых, за счет генерации силы, воздействующей на соседние актиновые филаменты, за счет циклирования миозиновых поперечных мостиков. Во-вторых, это перекачка ионов кальция из миоплазмы через саркоплазматический ретикулум против градиента их концентрации с использованием активного транспорта. Третьей функцией, которую выполняет АТФ, является активный транспорт ионов натрия и калия через сарколемму, так что ионы кальция могут высвобождаться при получении входного сигнала. Гидролиз АТФ управляет каждым из этих процессов.[16]
Гидролиз АТФ управляет каждым из этих процессов.[16]
Механизм
Многие процессы способны производить АТФ в организме, в зависимости от текущих метаболических условий. Производство АТФ может происходить в присутствии кислорода в результате клеточного дыхания, бета-окисления, кетоза, катаболизма липидов и белков, а также в анаэробных условиях.
Клеточное дыхание
Клеточное дыхание — это процесс катаболизма глюкозы в ацетил-КоА с образованием высокоэнергетических переносчиков электронов, которые будут окисляться во время окислительного фосфорилирования с образованием АТФ. Во время гликолиза, первой стадии клеточного дыхания, одна молекула глюкозы расщепляется на две молекулы пирувата. Во время этого процесса два АТФ продуцируются путем фосфорилирования субстрата ферментами PFK1 и пируваткиназой. Также происходит образование двух восстановленных молекул переносчика электронов NADH. Затем молекулы пирувата окисляются пируватдегидрогеназным комплексом с образованием молекулы ацетил-КоА. Затем молекула ацетил-КоА полностью окисляется с образованием диоксида углерода и восстановленных переносчиков электронов в цикле лимонной кислоты. При завершении цикла лимонной кислоты общий выход составляет две молекулы диоксида углерода, один эквивалент АТФ, три молекулы НАДН и одна молекула ФАДч3. Эти высокоэнергетические переносчики электронов затем переносят электроны в цепь переноса электронов, в которой ионы водорода (протоны) переносятся против своего градиента во внутреннее пространство мембраны из митохондриального матрикса. Затем молекулы АТФ синтезируются в виде протонов, движущихся вниз по электрохимическому градиенту мощности АТФ-синтазы.] Количество произведенного АТФ варьируется в зависимости от того, какой электронный носитель пожертвовал протоны. Одна молекула NADH производит две с половиной молекулы АТФ, тогда как одна молекула FADh3 производит полторы молекулы АТФ.[17]
Затем молекула ацетил-КоА полностью окисляется с образованием диоксида углерода и восстановленных переносчиков электронов в цикле лимонной кислоты. При завершении цикла лимонной кислоты общий выход составляет две молекулы диоксида углерода, один эквивалент АТФ, три молекулы НАДН и одна молекула ФАДч3. Эти высокоэнергетические переносчики электронов затем переносят электроны в цепь переноса электронов, в которой ионы водорода (протоны) переносятся против своего градиента во внутреннее пространство мембраны из митохондриального матрикса. Затем молекулы АТФ синтезируются в виде протонов, движущихся вниз по электрохимическому градиенту мощности АТФ-синтазы.] Количество произведенного АТФ варьируется в зависимости от того, какой электронный носитель пожертвовал протоны. Одна молекула NADH производит две с половиной молекулы АТФ, тогда как одна молекула FADh3 производит полторы молекулы АТФ.[17]
Бета-окисление
Бета-окисление — еще один механизм синтеза АТФ в организмах.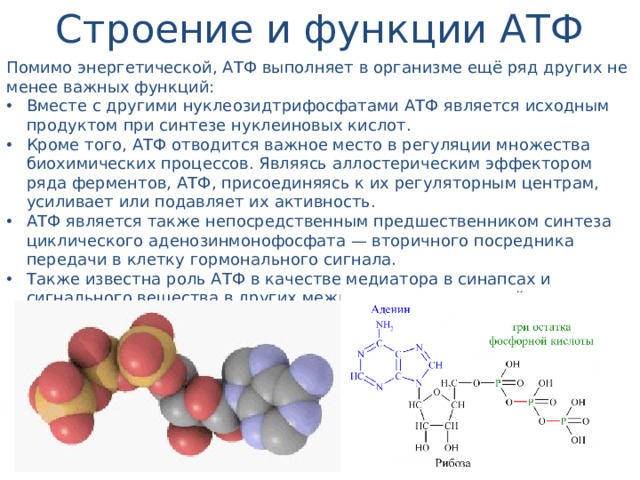 Во время бета-окисления цепи жирных кислот постоянно укорачиваются, образуя молекулы ацетил-КоА. На протяжении каждого цикла бета-окисления жирная кислота восстанавливается на две длины углерода, образуя одну молекулу ацетил-КоА, которая может быть окислена в цикле лимонной кислоты, и по одной молекуле НАДН и ФАДч3, которые передают свою высокую энергию электрон в транспортную цепь.[18]
Во время бета-окисления цепи жирных кислот постоянно укорачиваются, образуя молекулы ацетил-КоА. На протяжении каждого цикла бета-окисления жирная кислота восстанавливается на две длины углерода, образуя одну молекулу ацетил-КоА, которая может быть окислена в цикле лимонной кислоты, и по одной молекуле НАДН и ФАДч3, которые передают свою высокую энергию электрон в транспортную цепь.[18]
Кетоз
Кетоз – это реакция образования АТФ в результате катаболизма кетоновых тел. Во время кетоза кетоновые тела подвергаются катаболизму для производства энергии, образуя двадцать две молекулы АТФ и две молекулы ГТФ на молекулу ацетоацетата, которая окисляется в митохондриях.
Анаэробное дыхание
Когда кислорода мало или он недоступен во время клеточного дыхания, клетки могут подвергаться анаэробному дыханию. В анаэробных условиях происходит накопление молекул НАДН из-за неспособности окислять НАДН до НАД+, что ограничивает действие ГАФД и потребление глюкозы. Для поддержания гомеостатического уровня НАДН пируват восстанавливается до лактата, что приводит к окислению одной молекулы НАДН в процессе, известном как молочнокислое брожение. При молочнокислом брожении две молекулы НАДН, образующиеся в результате гликолиза, окисляются для поддержания резервуара НАД+. Эта реакция дает только две молекулы АТФ на молекулу глюкозы.
Для поддержания гомеостатического уровня НАДН пируват восстанавливается до лактата, что приводит к окислению одной молекулы НАДН в процессе, известном как молочнокислое брожение. При молочнокислом брожении две молекулы НАДН, образующиеся в результате гликолиза, окисляются для поддержания резервуара НАД+. Эта реакция дает только две молекулы АТФ на молекулу глюкозы.
Связанное тестирование
Многие методы позволяют рассчитать внутриклеточный уровень АТФ. Общепринятый протокол включает использование люциферазы светлячка, фермента, который вызывает окисление люциферина.[19] Эта реакция поддается количественному измерению из-за выхода энергии этой реакции, высвобождающей фотон света, известного как биолюминесценция, которая поддается количественному измерению.
Клиническое значение
Роль АТФ в контроле боли
ATP демонстрирует уменьшение острой периоперационной боли в клинических исследованиях.[20] В этих исследованиях пациенты получали АТФ внутривенно.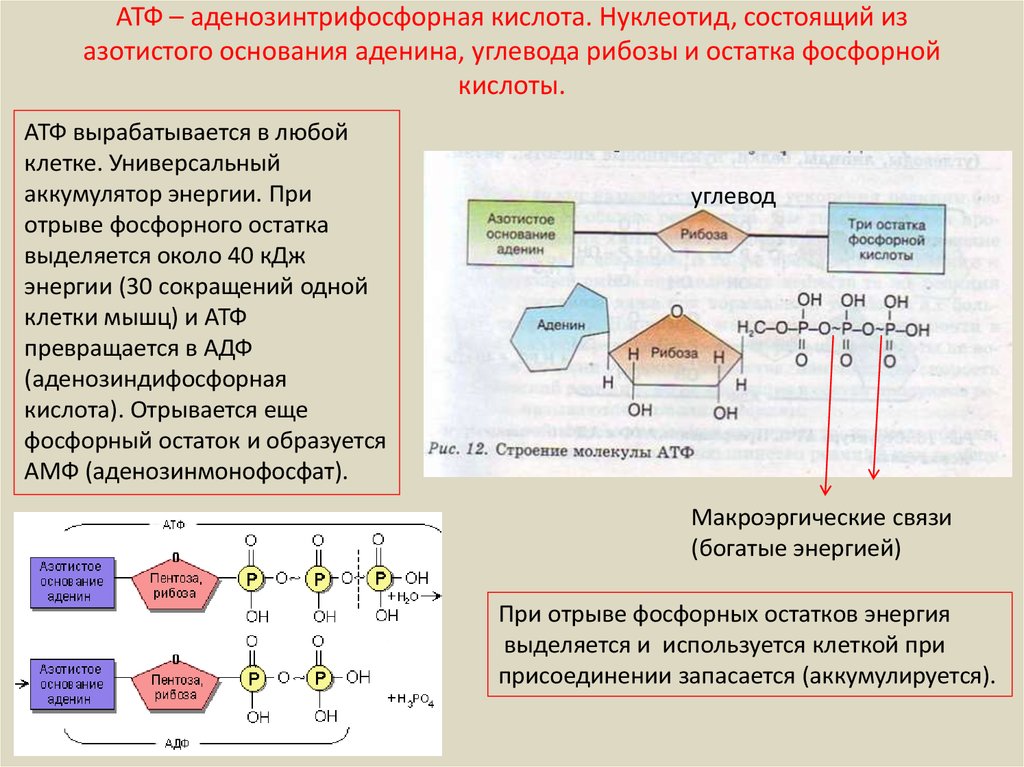 Внутривенная инфузия аденозина действует на аденозиновый рецептор A1, инициируя сигнальный каскад, который в конечном итоге способствует обезболивающему эффекту, наблюдаемому при воспалении. Исследования показали, что соединения аденозина уменьшают аллодинию и гипералгезию при введении в умеренных дозах.[20] Активация аденозинового рецептора A1 оказывает эффективное обезболивающее действие благодаря медленному началу и длительному действию, которое в некоторых случаях может продолжаться в течение нескольких недель.
Внутривенная инфузия аденозина действует на аденозиновый рецептор A1, инициируя сигнальный каскад, который в конечном итоге способствует обезболивающему эффекту, наблюдаемому при воспалении. Исследования показали, что соединения аденозина уменьшают аллодинию и гипералгезию при введении в умеренных дозах.[20] Активация аденозинового рецептора A1 оказывает эффективное обезболивающее действие благодаря медленному началу и длительному действию, которое в некоторых случаях может продолжаться в течение нескольких недель.
Анестезия
Добавление АТФ дало положительные результаты во время анестезии. Имеющиеся данные показывают, что низкие дозы аденозина уменьшают невропатическую боль, ишемическую боль и гипералгезию до уровня, сравнимого с морфином.[21] Аденозин также снижал послеоперационное использование опиоидов, что свидетельствует о потенциальной длительной активации аденозиновых рецепторов А1.
Кардиология и хирургия
Было показано, что АТФ является безопасным и практичным легочным сосудорасширяющим средством у пациентов с легочной гипертензией. [21] Точно так же аденозин и АТФ можно использовать во время операции, чтобы вызвать гипотензию у пациентов.[21]
[21] Точно так же аденозин и АТФ можно использовать во время операции, чтобы вызвать гипотензию у пациентов.[21]
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Литература
- 1.
Meurer F, Do HT, Sadowski G, Held C. Стандартная энергия Гиббса метаболических реакций: II. Глюкозо-6-фосфатазная реакция и гидролиз АТФ. Биофиз хим. 2017 Апр; 223:30-38. [PubMed: 28282626]
- 2.
Beis I, Newsholme EA. Содержание адениновых нуклеотидов, фосфагенов и некоторых гликолитических интермедиатов в покоящихся мышцах позвоночных и беспозвоночных. Биохим Дж. 1975 октября; 152(1):23-32. [Бесплатная статья PMC: PMC1172435] [PubMed: 1212224]
- 3.
Wang X, Zhang X, Wu D, Huang Z, Hou T, Jian C, Yu P, Lu F, Zhang R, Sun T, Li J, Qi W, Wang Y, Gao F, Cheng H. Митохондриальные вспышки регулируют гомеостаз АТФ в сердце.
 Элиф. 2017 Jul 10;6 [PMC бесплатная статья: PMC5503511] [PubMed: 28692422]
Элиф. 2017 Jul 10;6 [PMC бесплатная статья: PMC5503511] [PubMed: 28692422]- 4.
Mishra NS, Tuteja R, Tuteja N. Передача сигналов через сети киназ MAP в растениях. Арх Биохим Биофиз. 2006 01 августа; 452 (1): 55-68. [В паблике: 16806044]
- 5.
Лин Х, Айрапетов М.К., Сун Г. Характеристика взаимодействий между активным центром протеинтирозинкиназы и активатором двухвалентного металла. БМС Биохим. 2005 23 ноября; 6:25. [PMC бесплатная статья: PMC1316873] [PubMed: 16305747]
- 6.
Zimmermann H. Внеклеточный АТФ и другие нуклеотиды-повсеместные триггеры высвобождения межклеточного мессенджера. Пуринергический сигнал. 2016 март; 12(1):25-57. [Бесплатная статья PMC: PMC4749530] [PubMed: 26545760]
- 7.
Каменецкий М., Миддельхауфе С., Банк Э.М., Левин Л.Р., Бак Дж., Стигборн С. Молекулярные детали образования цАМФ в клетках млекопитающих: рассказ о двух системах. Дж Мол Биол. 2006 г., 29 сентября; 362(4):623-39.
 [Бесплатная статья PMC: PMC3662476] [PubMed: 16934836]
[Бесплатная статья PMC: PMC3662476] [PubMed: 16934836]- 8.
Джойс CM, Steitz TA. Структуры и функции полимеразы: вариации на тему? J Бактериол. 1995 ноябрь; 177 (22): 6321-9. [Бесплатная статья PMC: PMC177480] [PubMed: 7592405]
- 9.
Bonora M, Patergnani S, Rimessi A, De Marchi E, Suski JM, Bononi A, Giorgi C, Marchi S, Missiroli S, Poletti F, Wieckowski MR, Pinton P. Синтез и хранение АТФ. Пуринергический сигнал. 2012 Сентябрь;8(3):343-57. [Бесплатная статья PMC: PMC3360099] [PubMed: 22528680]
- 10.
Карденас С., Миллер Р.А., Смит И., Буй Т., Молго Дж., Мюллер М., Вайс Х., Чунг К.Х., Ян Дж., Паркер И., Томпсон С.Б., Бирнбаум М.Дж., Халлоус К.Р., Фоскетт Дж.К. Существенная регуляция биоэнергетики клетки путем конститутивного переноса Ca2+ рецептора InsP3 в митохондрии. Клетка. 2010 23 июля; 142 (2): 270-83. [Бесплатная статья PMC: PMC2911450] [PubMed: 20655468]
- 11.
Пабло Хуидобро-Торо Дж.
 , Вероника Доносо М. Симпатическая котрансмиссия: скоординированное действие АТФ и норадреналина и их модуляция нейропептидом Y в сосудистых нейроэффекторных соединениях человека. Евр Дж Фармакол. 2004 01 октября; 500 (1-3): 27-35. [PubMed: 15464018]
, Вероника Доносо М. Симпатическая котрансмиссия: скоординированное действие АТФ и норадреналина и их модуляция нейропептидом Y в сосудистых нейроэффекторных соединениях человека. Евр Дж Фармакол. 2004 01 октября; 500 (1-3): 27-35. [PubMed: 15464018]- 12.
Coco S, Calegari F, Pravettoni E, Pozzi D, Taverna E, Rosa P, Matteoli M, Verderio C. Хранение и высвобождение АТФ из астроцитов в культуре. Дж. Биол. Хим. 2003 10 января; 278 (2): 1354-62. [В паблике: 12414798]
- 13.
Аттвелл Д., Лафлин С.Б. Энергетический баланс для передачи сигналов в сером веществе головного мозга. J Cereb Blood Flow Metab. 21 октября 2001 г. (10): 1133-45. [PubMed: 11598490]
- 14.
Harris JJ, Jolivet R, Attwell D. Использование и поставка синаптической энергии. Нейрон. 06 сентября 2012 г .; 75 (5): 762–777. [PubMed: 22958818]
- 15.
Вонг-Райли MT. Цитохромоксидаза: эндогенный метаболический маркер активности нейронов.
 Тренды Нейроси. 1989 марта; 12(3):94-101. [PubMed: 2469224]
Тренды Нейроси. 1989 марта; 12(3):94-101. [PubMed: 2469224]- 16.
Барклай CJ. Энергетика сокращения. сост. физиол. 2015 Апрель;5(2):961-95. [PubMed: 25880520]
- 17.
Богатый PR. Молекулярный механизм дыхательной цепи Кейлина. Биохим Сок Транс. 2003 г.; 31 декабря (часть 6): 1095–105. [PubMed: 14641005]
- 18.
Ronnett GV, Kim EK, Landree LE, Tu Y. Метаболизм жирных кислот как мишень для лечения ожирения. Физиол Поведение. 2005 19 мая;85(1):25-35. [PubMed: 15878185]
- 19.
Бровко Л.Ю., Романова Н.А., Угарова Н.Н. Биолюминесцентный анализ бактериального внутриклеточного АМФ, АДФ и АТФ с использованием коиммобилизованного трехферментного реагента (аденилаткиназа, пируваткиназа и люцифераза светлячка). Анальная биохимия. 1994 01 августа; 220 (2): 410-4. [PubMed: 7978286]
- 20.
Хаяшида М., Фукуда К., Фукунага А. Клиническое применение аденозина и АТФ для обезболивания.
 Джей Анест. 2005;19(3): 225-35. [PubMed: 16032451]
Джей Анест. 2005;19(3): 225-35. [PubMed: 16032451]- 21.
Agteresch HJ, Dagnelie PC, van den Berg JW, Wilson JH. Аденозинтрифосфат: установившееся и потенциальное клиническое применение. Наркотики. 1999 авг; 58 (2): 211-32. [PubMed: 10473017]
Физиология, Аденозинтрифосфат — StatPearls
Введение
Тело представляет собой сложный организм, и поэтому ему требуется энергия для поддержания нормального функционирования. Аденозинтрифосфат (АТФ) является источником энергии для использования и хранения на клеточном уровне. Структура АТФ представляет собой нуклеозидтрифосфат, состоящий из азотистого основания (аденина), сахара рибозы и трех последовательно связанных фосфатных групп. АТФ обычно называют «энергетической валютой» клетки, поскольку он обеспечивает легко высвобождаемую энергию в связи между второй и третьей фосфатными группами. Помимо обеспечения энергией, расщепление АТФ посредством гидролиза выполняет широкий спектр клеточных функций, включая передачу сигналов и синтез ДНК/РНК. Синтез АТФ использует энергию, полученную от нескольких катаболических механизмов, включая клеточное дыхание, бета-окисление и кетоз.
Синтез АТФ использует энергию, полученную от нескольких катаболических механизмов, включая клеточное дыхание, бета-окисление и кетоз.
Большая часть синтеза АТФ происходит при клеточном дыхании в митохондриальном матриксе: образуется примерно тридцать две молекулы АТФ на молекулу окисляемой глюкозы. АТФ потребляется для получения энергии в процессах, включая перенос ионов, сокращение мышц, распространение нервных импульсов, фосфорилирование субстрата и химический синтез. Эти процессы, как и другие, создают высокий спрос на АТФ. В результате клетки в организме человека зависят от гидролиза от 100 до 150 моль АТФ в день для обеспечения правильного функционирования. В следующих разделах будет проведена дальнейшая оценка роли АТФ как ключевой молекулы в ежедневном функционировании клетки.
Сотовый уровень
АТФ является отличным накопителем энергии для использования в качестве «валюты» благодаря фосфатным группам, которые соединяются посредством фосфодиэфирных связей. Эти связи имеют высокую энергию из-за связанных с ними электроотрицательных зарядов, создающих силу отталкивания между фосфатными группами. Значительное количество энергии остается запасенным в фосфатно-фосфатных связях. В ходе метаболических процессов АТФ гидролизуется до АДФ или далее до АМФ и свободных неорганических фосфатных групп. Процесс гидролиза АТФ до АДФ энергетически выгоден, давая свободную энергию Гиббса -7,3 кал/моль.[1] АТФ должен постоянно пополняться, чтобы питать постоянно работающую клетку. Обычная внутриклеточная концентрация АТФ составляет от 1 до 10 мкМ.[2] Существует множество механизмов обратной связи, обеспечивающих поддержание постоянного уровня АТФ в клетке. Усиление или ингибирование АТФ-синтазы является обычным регуляторным механизмом. Например, АТФ ингибирует фосфофруктокиназу-1 (PFK1) и пируваткиназу, два ключевых фермента гликолиза, эффективно действуя как петля отрицательной обратной связи для ингибирования распада глюкозы при наличии достаточного количества клеточного АТФ.
Эти связи имеют высокую энергию из-за связанных с ними электроотрицательных зарядов, создающих силу отталкивания между фосфатными группами. Значительное количество энергии остается запасенным в фосфатно-фосфатных связях. В ходе метаболических процессов АТФ гидролизуется до АДФ или далее до АМФ и свободных неорганических фосфатных групп. Процесс гидролиза АТФ до АДФ энергетически выгоден, давая свободную энергию Гиббса -7,3 кал/моль.[1] АТФ должен постоянно пополняться, чтобы питать постоянно работающую клетку. Обычная внутриклеточная концентрация АТФ составляет от 1 до 10 мкМ.[2] Существует множество механизмов обратной связи, обеспечивающих поддержание постоянного уровня АТФ в клетке. Усиление или ингибирование АТФ-синтазы является обычным регуляторным механизмом. Например, АТФ ингибирует фосфофруктокиназу-1 (PFK1) и пируваткиназу, два ключевых фермента гликолиза, эффективно действуя как петля отрицательной обратной связи для ингибирования распада глюкозы при наличии достаточного количества клеточного АТФ.
И наоборот, АДФ и АМФ могут активировать PFK1 и пируваткиназу, способствуя синтезу АТФ в периоды высокой потребности в энергии. Другие системы регулируют АТФ, например, в регуляторных механизмах, участвующих в регуляции синтеза АТФ в сердце. Новые эксперименты показали, что десятисекундные всплески, называемые митохондриальными вспышками, могут нарушить выработку АТФ в сердце. Во время этих митохондриальных вспышек митохондрии выделяют активные формы кислорода и эффективно приостанавливают синтез АТФ. Ингибирование продукции АТФ происходит во время митохондриальных вспышек. Во время низкой потребности в энергии, когда клетки сердечной мышцы получали достаточно строительных блоков, необходимых для производства АТФ, митохондриальные вспышки наблюдались чаще. С другой стороны, когда потребность в энергии высока во время быстрого сердечного сокращения, митохондриальные вспышки происходили реже. Эти результаты свидетельствуют о том, что в периоды, когда необходимы значительные количества АТФ, митохондриальные вспышки происходят реже, чтобы обеспечить продолжение производства АТФ.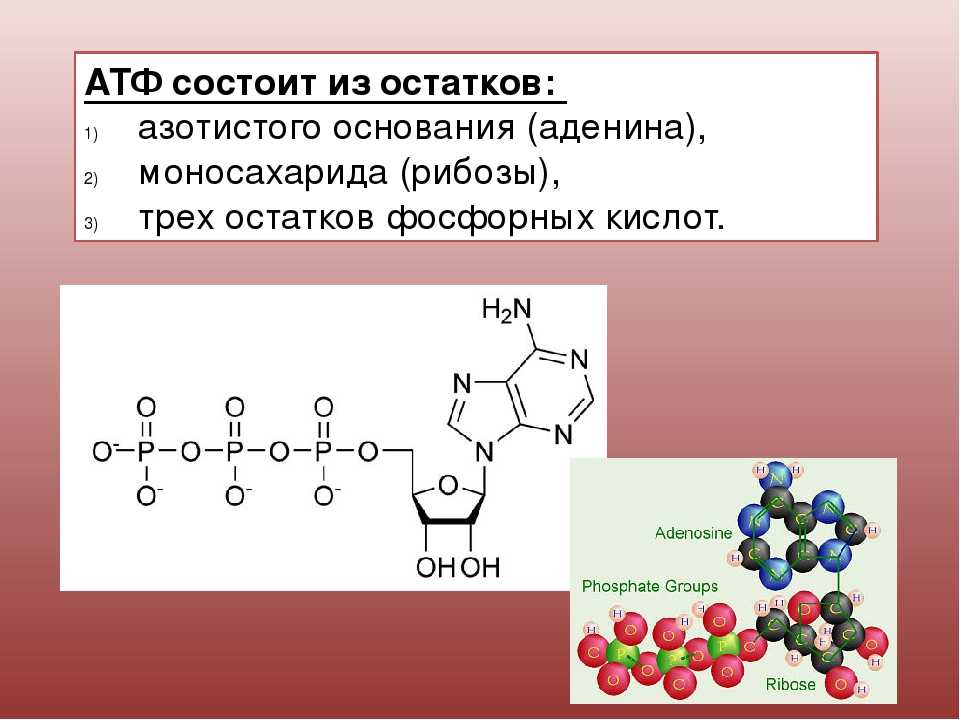 И наоборот, в периоды низкой выработки энергии митохондриальные вспышки происходили более регулярно и ингибировали выработку АТФ.[3]
И наоборот, в периоды низкой выработки энергии митохондриальные вспышки происходили более регулярно и ингибировали выработку АТФ.[3]
Функция
Гидролиз АТФ обеспечивает энергию, необходимую для многих важных процессов в организмах и клетках. К ним относятся внутриклеточная передача сигналов, синтез ДНК и РНК, пуринергическая передача сигналов, синаптическая передача сигналов, активный транспорт и сокращение мышц. Эти темы не являются исчерпывающим списком, но включают в себя некоторые жизненно важные роли, которые выполняет ATP.
АТФ во внутриклеточной передаче сигналов
Передача сигнала в значительной степени зависит от АТФ. АТФ может служить субстратом для киназ, самого многочисленного АТФ-связывающего белка. Когда киназа фосфорилирует белок, может активироваться сигнальный каскад, что приводит к модуляции различных внутриклеточных сигнальных путей.[4] Активность киназы жизненно важна для клетки и, следовательно, должна жестко регулироваться.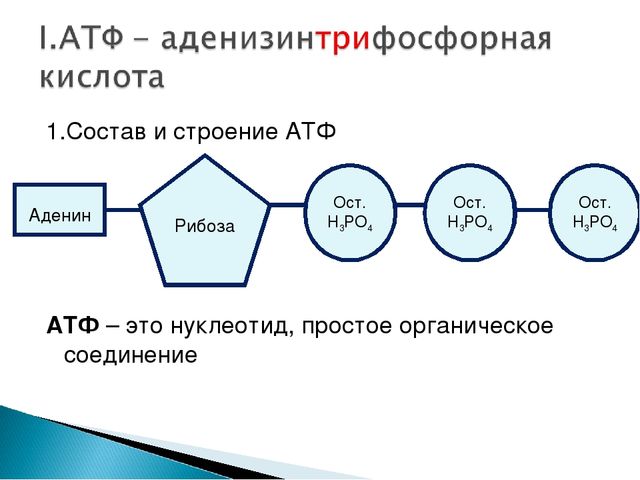 Присутствие иона магния помогает регулировать активность киназы.[5] Регуляция осуществляется через ионы магния, находящиеся в клетке в виде комплекса с АТФ, связанного на фосфатно-кислородных центрах. В дополнение к киназной активности АТФ может функционировать как повсеместный триггер высвобождения внутриклеточного мессенджера.[6] Эти мессенджеры включают гормоны, различные ферменты, липидные медиаторы, нейротрансмиттеры, оксид азота, факторы роста и активные формы кислорода.[6] Пример использования АТФ во внутриклеточной передаче сигналов можно наблюдать в АТФ, выступающем в качестве субстрата для аденилатциклазы. Этот процесс в основном происходит в сигнальных путях рецепторов, связанных с G-белком. При связывании с аденилатциклазой АТФ превращается в циклический АМФ, который способствует высвобождению кальция из внутриклеточных запасов.[7] У цАМФ есть и другие роли, в том числе вторичные мессенджеры в сигнальных каскадах гормонов, активация протеинкиназ и регуляция функции ионных каналов.
Присутствие иона магния помогает регулировать активность киназы.[5] Регуляция осуществляется через ионы магния, находящиеся в клетке в виде комплекса с АТФ, связанного на фосфатно-кислородных центрах. В дополнение к киназной активности АТФ может функционировать как повсеместный триггер высвобождения внутриклеточного мессенджера.[6] Эти мессенджеры включают гормоны, различные ферменты, липидные медиаторы, нейротрансмиттеры, оксид азота, факторы роста и активные формы кислорода.[6] Пример использования АТФ во внутриклеточной передаче сигналов можно наблюдать в АТФ, выступающем в качестве субстрата для аденилатциклазы. Этот процесс в основном происходит в сигнальных путях рецепторов, связанных с G-белком. При связывании с аденилатциклазой АТФ превращается в циклический АМФ, который способствует высвобождению кальция из внутриклеточных запасов.[7] У цАМФ есть и другие роли, в том числе вторичные мессенджеры в сигнальных каскадах гормонов, активация протеинкиназ и регуляция функции ионных каналов.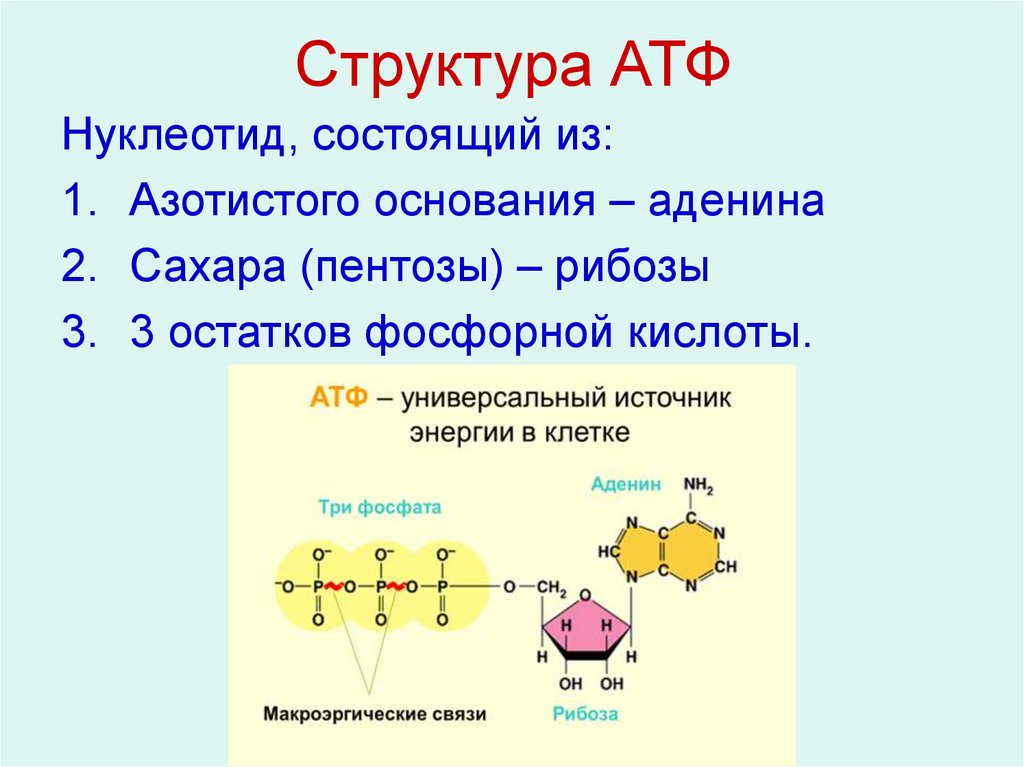
Синтез ДНК/РНК
Для синтеза ДНК и РНК требуется АТФ. АТФ является одним из четырех нуклеотид-трифосфатных мономеров, необходимых при синтезе РНК. Синтез ДНК использует аналогичный механизм, за исключением того, что при синтезе ДНК АТФ сначала трансформируется путем удаления атома кислорода из сахара с образованием дезоксирибонуклеотида, дАТФ.
Пуринергическая сигнализация
Пуринергическая передача сигналов представляет собой форму внеклеточной паракринной передачи сигналов, которая опосредована пуриновыми нуклеотидами, включая АТФ. Этот процесс обычно влечет за собой активацию пуринергических рецепторов на клетках в непосредственной близости, тем самым передавая сигналы для регулирования внутриклеточных процессов. АТФ высвобождается из везикулярных запасов и регулируется IP3 и другими распространенными экзоцитотическими регуляторными механизмами. АТФ хранится и высвобождается вместе с нейротрансмиттерами, что еще раз подтверждает представление о том, что АТФ является необходимым медиатором пуринергической нейротрансмиссии как в симпатических, так и в парасимпатических нервах.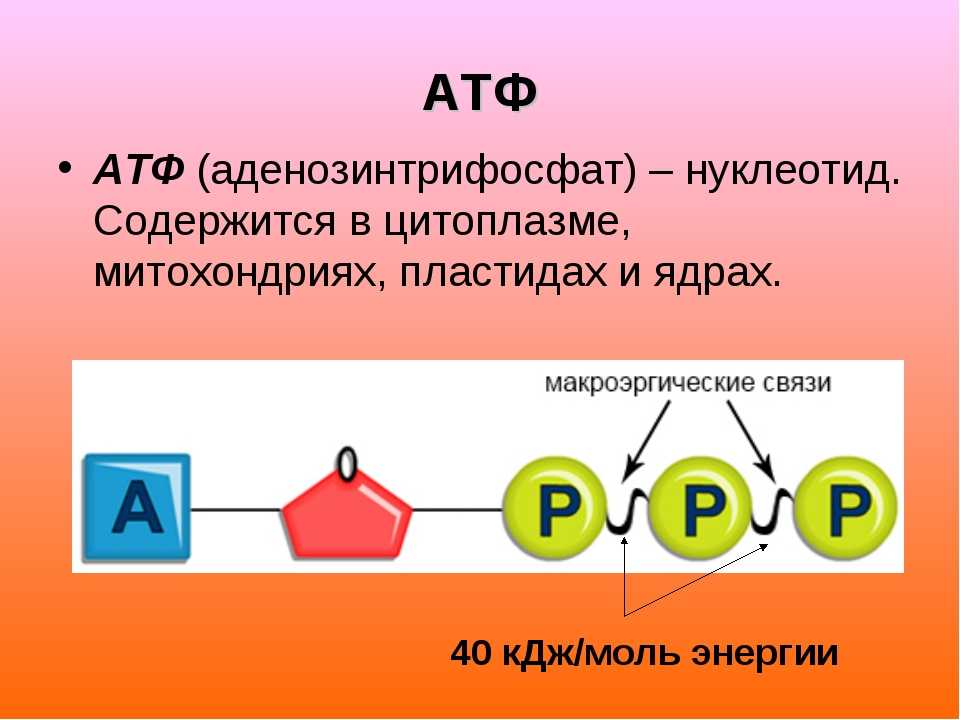 АТФ может индуцировать несколько пуринергических реакций, включая контроль вегетативных функций, взаимодействие нервной глии, боль и контроль тонуса сосудов.[9]][10][11][12]
АТФ может индуцировать несколько пуринергических реакций, включая контроль вегетативных функций, взаимодействие нервной глии, боль и контроль тонуса сосудов.[9]][10][11][12]
Нейротрансмиссия
Мозг является самым большим потребителем АТФ в организме, потребляя примерно двадцать пять процентов всей доступной энергии.[13] Большое количество энергии тратится на поддержание концентрации ионов для правильной передачи сигналов нейронами и синаптической передачи.[14] Синаптическая передача является энергоемким процессом. На пресинаптических окончаниях АТФ требуется для установления ионного градиента, который перемещает нейротрансмиттеры в везикулы, и для подготовки везикул к высвобождению посредством экзоцитоза. [14] Нейрональная передача сигналов зависит от потенциала действия, достигающего пресинаптических окончаний, сигнализирующего о высвобождении загруженных везикул. Этот процесс зависит от того, что АТФ восстанавливает концентрацию ионов в аксоне после каждого потенциала действия, позволяя возникать другому сигналу.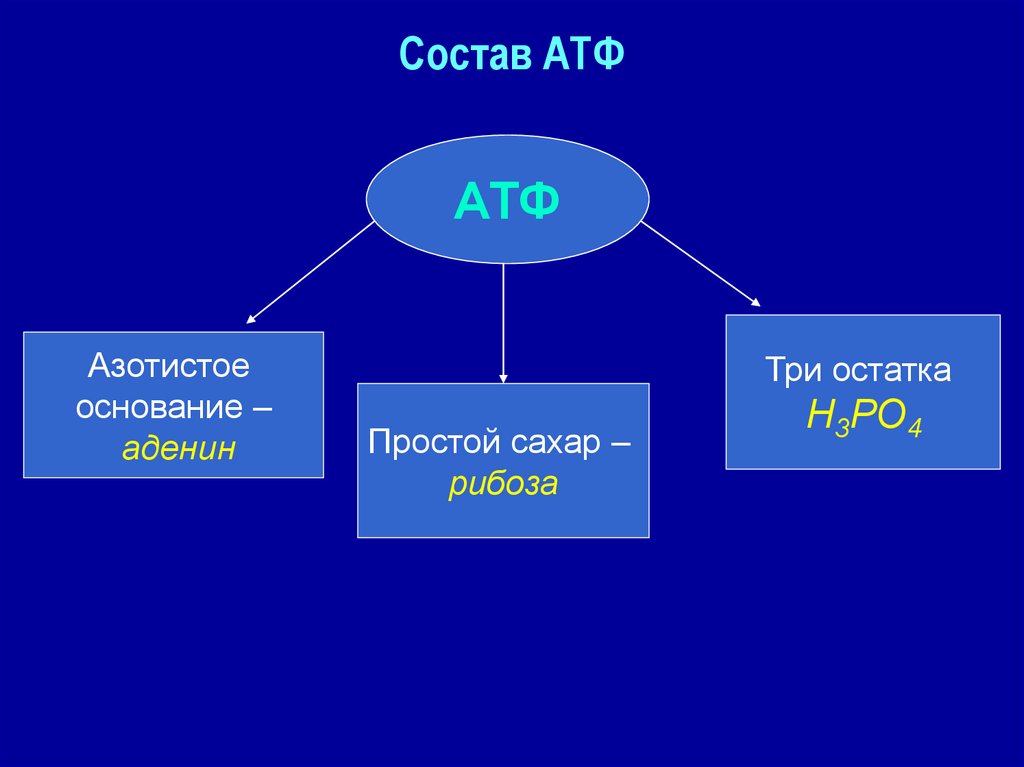 Активный транспорт отвечает за сброс концентраций ионов натрия и калия до исходных значений после возникновения потенциала действия через Na/K-АТФазу. Во время этого процесса одна молекула АТФ гидролизуется, три иона натрия транспортируются из клетки и два иона калия транспортируются обратно в клетку, оба из которых движутся против градиента их концентрации.
Активный транспорт отвечает за сброс концентраций ионов натрия и калия до исходных значений после возникновения потенциала действия через Na/K-АТФазу. Во время этого процесса одна молекула АТФ гидролизуется, три иона натрия транспортируются из клетки и два иона калия транспортируются обратно в клетку, оба из которых движутся против градиента их концентрации.
Потенциалы действия, проходящие по аксону, инициируют везикулярное высвобождение при достижении пресинаптического окончания. После установления ионного градиента потенциалы действия затем распространяются вниз по аксону посредством деполяризации аксона, посылая сигнал к терминалу. Приблизительно один миллиард ионов натрия необходим для распространения одного потенциала действия. Нейронам потребуется гидролизовать около миллиарда молекул АТФ, чтобы восстановить концентрацию ионов натрия/калия после каждой клеточной деполяризации.[13]Возбуждающие синапсы в значительной степени преобладают в сером веществе мозга. Везикулы, содержащие глутамат, высвобождаются в синаптическую щель, чтобы активировать постсинаптические возбуждающие глутаминергические рецепторы.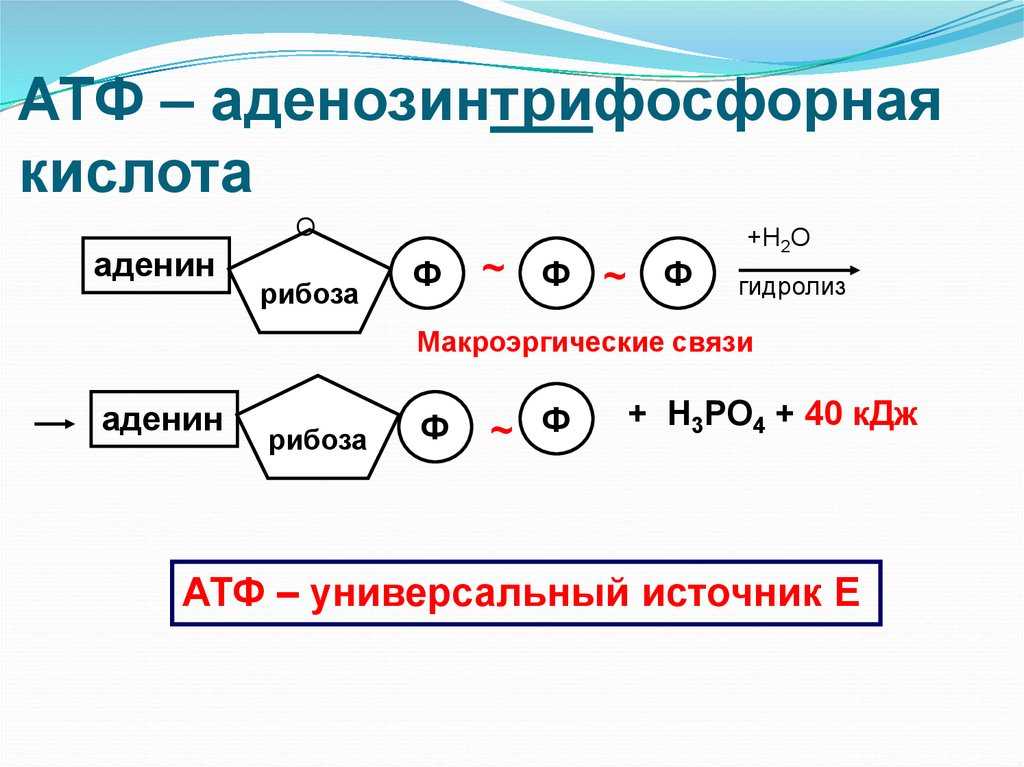 Для загрузки этих молекул требуется большое количество АТФ из-за почти четырех тысяч молекул глутамата, хранящихся в одном пузырьке. Необходимы значительные запасы энергии, чтобы инициировать высвобождение везикул, управлять глутаматергическими постсинаптическими процессами и перерабатывать везикулы, а также оставшийся глутамат. Следовательно, из-за большого количества энергии, необходимой для упаковки глутамата, митохондрии близки к глутаматергическим пузырькам.
Для загрузки этих молекул требуется большое количество АТФ из-за почти четырех тысяч молекул глутамата, хранящихся в одном пузырьке. Необходимы значительные запасы энергии, чтобы инициировать высвобождение везикул, управлять глутаматергическими постсинаптическими процессами и перерабатывать везикулы, а также оставшийся глутамат. Следовательно, из-за большого количества энергии, необходимой для упаковки глутамата, митохондрии близки к глутаматергическим пузырькам.
АТФ в сокращении мышц
Сокращение мышц является необходимой функцией повседневной жизни и не может происходить без АТФ. Есть три основные роли, которые АТФ выполняет в действии мышечного сокращения. Во-первых, за счет генерации силы, воздействующей на соседние актиновые филаменты, за счет циклирования миозиновых поперечных мостиков. Во-вторых, это перекачка ионов кальция из миоплазмы через саркоплазматический ретикулум против градиента их концентрации с использованием активного транспорта. Третьей функцией, которую выполняет АТФ, является активный транспорт ионов натрия и калия через сарколемму, так что ионы кальция могут высвобождаться при получении входного сигнала. Гидролиз АТФ управляет каждым из этих процессов.[16]
Гидролиз АТФ управляет каждым из этих процессов.[16]
Механизм
Многие процессы способны производить АТФ в организме, в зависимости от текущих метаболических условий. Производство АТФ может происходить в присутствии кислорода в результате клеточного дыхания, бета-окисления, кетоза, катаболизма липидов и белков, а также в анаэробных условиях.
Клеточное дыхание
Клеточное дыхание — это процесс катаболизма глюкозы в ацетил-КоА с образованием высокоэнергетических переносчиков электронов, которые будут окисляться во время окислительного фосфорилирования с образованием АТФ. Во время гликолиза, первой стадии клеточного дыхания, одна молекула глюкозы расщепляется на две молекулы пирувата. Во время этого процесса два АТФ продуцируются путем фосфорилирования субстрата ферментами PFK1 и пируваткиназой. Также происходит образование двух восстановленных молекул переносчика электронов NADH. Затем молекулы пирувата окисляются пируватдегидрогеназным комплексом с образованием молекулы ацетил-КоА.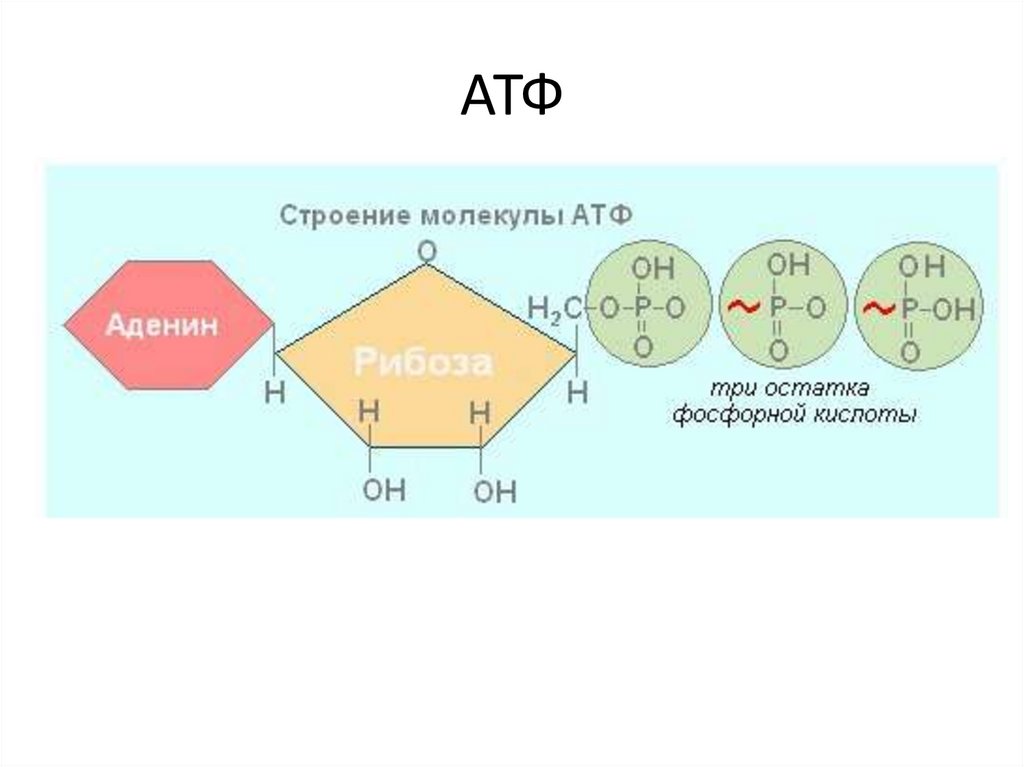 Затем молекула ацетил-КоА полностью окисляется с образованием диоксида углерода и восстановленных переносчиков электронов в цикле лимонной кислоты. При завершении цикла лимонной кислоты общий выход составляет две молекулы диоксида углерода, один эквивалент АТФ, три молекулы НАДН и одна молекула ФАДч3. Эти высокоэнергетические переносчики электронов затем переносят электроны в цепь переноса электронов, в которой ионы водорода (протоны) переносятся против своего градиента во внутреннее пространство мембраны из митохондриального матрикса. Затем молекулы АТФ синтезируются в виде протонов, движущихся вниз по электрохимическому градиенту мощности АТФ-синтазы.] Количество произведенного АТФ варьируется в зависимости от того, какой электронный носитель пожертвовал протоны. Одна молекула NADH производит две с половиной молекулы АТФ, тогда как одна молекула FADh3 производит полторы молекулы АТФ.[17]
Затем молекула ацетил-КоА полностью окисляется с образованием диоксида углерода и восстановленных переносчиков электронов в цикле лимонной кислоты. При завершении цикла лимонной кислоты общий выход составляет две молекулы диоксида углерода, один эквивалент АТФ, три молекулы НАДН и одна молекула ФАДч3. Эти высокоэнергетические переносчики электронов затем переносят электроны в цепь переноса электронов, в которой ионы водорода (протоны) переносятся против своего градиента во внутреннее пространство мембраны из митохондриального матрикса. Затем молекулы АТФ синтезируются в виде протонов, движущихся вниз по электрохимическому градиенту мощности АТФ-синтазы.] Количество произведенного АТФ варьируется в зависимости от того, какой электронный носитель пожертвовал протоны. Одна молекула NADH производит две с половиной молекулы АТФ, тогда как одна молекула FADh3 производит полторы молекулы АТФ.[17]
Бета-окисление
Бета-окисление — еще один механизм синтеза АТФ в организмах.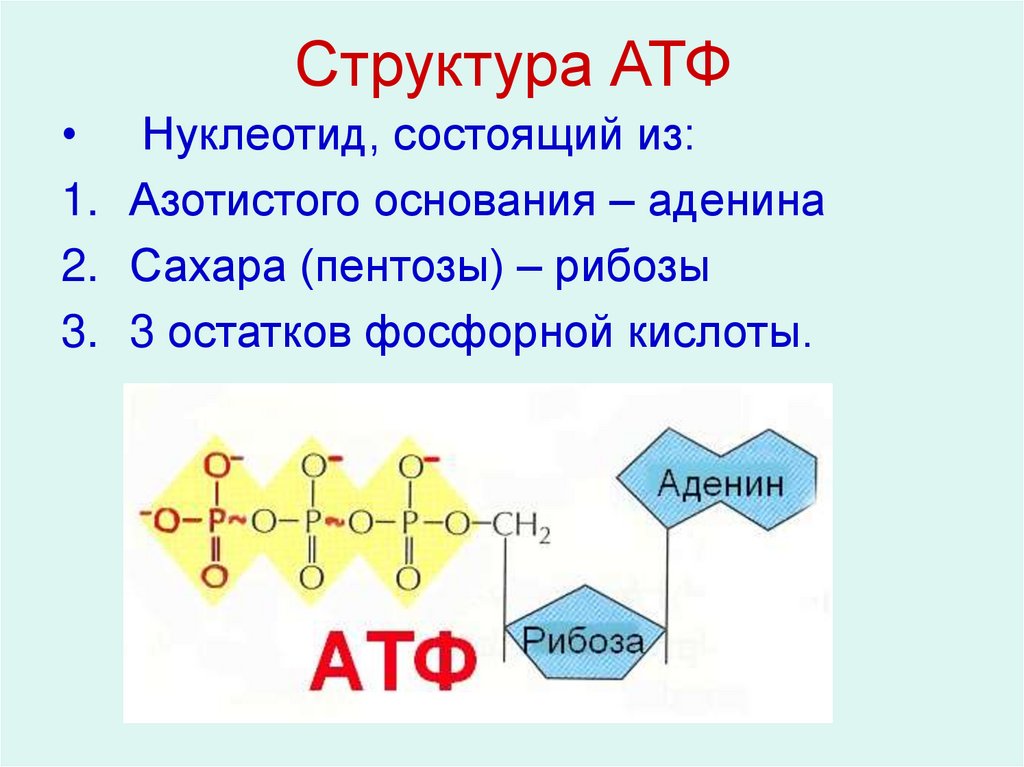 Во время бета-окисления цепи жирных кислот постоянно укорачиваются, образуя молекулы ацетил-КоА. На протяжении каждого цикла бета-окисления жирная кислота восстанавливается на две длины углерода, образуя одну молекулу ацетил-КоА, которая может быть окислена в цикле лимонной кислоты, и по одной молекуле НАДН и ФАДч3, которые передают свою высокую энергию электрон в транспортную цепь.[18]
Во время бета-окисления цепи жирных кислот постоянно укорачиваются, образуя молекулы ацетил-КоА. На протяжении каждого цикла бета-окисления жирная кислота восстанавливается на две длины углерода, образуя одну молекулу ацетил-КоА, которая может быть окислена в цикле лимонной кислоты, и по одной молекуле НАДН и ФАДч3, которые передают свою высокую энергию электрон в транспортную цепь.[18]
Кетоз
Кетоз – это реакция образования АТФ в результате катаболизма кетоновых тел. Во время кетоза кетоновые тела подвергаются катаболизму для производства энергии, образуя двадцать две молекулы АТФ и две молекулы ГТФ на молекулу ацетоацетата, которая окисляется в митохондриях.
Анаэробное дыхание
Когда кислорода мало или он недоступен во время клеточного дыхания, клетки могут подвергаться анаэробному дыханию. В анаэробных условиях происходит накопление молекул НАДН из-за неспособности окислять НАДН до НАД+, что ограничивает действие ГАФД и потребление глюкозы. Для поддержания гомеостатического уровня НАДН пируват восстанавливается до лактата, что приводит к окислению одной молекулы НАДН в процессе, известном как молочнокислое брожение. При молочнокислом брожении две молекулы НАДН, образующиеся в результате гликолиза, окисляются для поддержания резервуара НАД+. Эта реакция дает только две молекулы АТФ на молекулу глюкозы.
Для поддержания гомеостатического уровня НАДН пируват восстанавливается до лактата, что приводит к окислению одной молекулы НАДН в процессе, известном как молочнокислое брожение. При молочнокислом брожении две молекулы НАДН, образующиеся в результате гликолиза, окисляются для поддержания резервуара НАД+. Эта реакция дает только две молекулы АТФ на молекулу глюкозы.
Связанное тестирование
Многие методы позволяют рассчитать внутриклеточный уровень АТФ. Общепринятый протокол включает использование люциферазы светлячка, фермента, который вызывает окисление люциферина.[19] Эта реакция поддается количественному измерению из-за выхода энергии этой реакции, высвобождающей фотон света, известного как биолюминесценция, которая поддается количественному измерению.
Клиническое значение
Роль АТФ в контроле боли
ATP демонстрирует уменьшение острой периоперационной боли в клинических исследованиях.[20] В этих исследованиях пациенты получали АТФ внутривенно.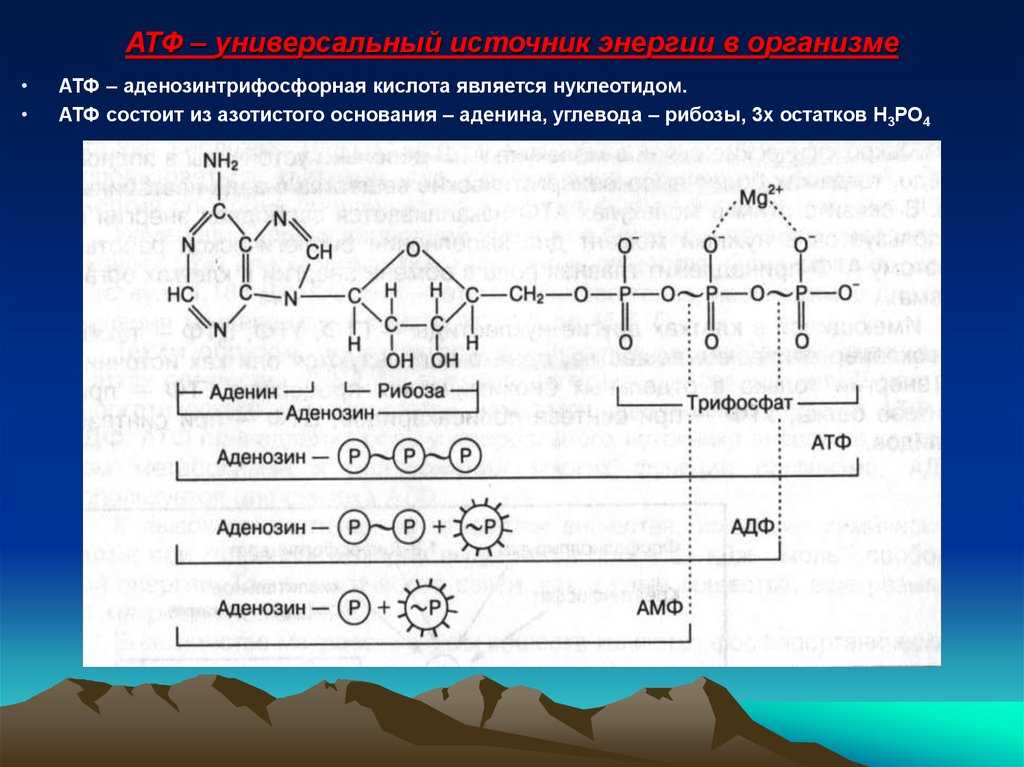 Внутривенная инфузия аденозина действует на аденозиновый рецептор A1, инициируя сигнальный каскад, который в конечном итоге способствует обезболивающему эффекту, наблюдаемому при воспалении. Исследования показали, что соединения аденозина уменьшают аллодинию и гипералгезию при введении в умеренных дозах.[20] Активация аденозинового рецептора A1 оказывает эффективное обезболивающее действие благодаря медленному началу и длительному действию, которое в некоторых случаях может продолжаться в течение нескольких недель.
Внутривенная инфузия аденозина действует на аденозиновый рецептор A1, инициируя сигнальный каскад, который в конечном итоге способствует обезболивающему эффекту, наблюдаемому при воспалении. Исследования показали, что соединения аденозина уменьшают аллодинию и гипералгезию при введении в умеренных дозах.[20] Активация аденозинового рецептора A1 оказывает эффективное обезболивающее действие благодаря медленному началу и длительному действию, которое в некоторых случаях может продолжаться в течение нескольких недель.
Анестезия
Добавление АТФ дало положительные результаты во время анестезии. Имеющиеся данные показывают, что низкие дозы аденозина уменьшают невропатическую боль, ишемическую боль и гипералгезию до уровня, сравнимого с морфином.[21] Аденозин также снижал послеоперационное использование опиоидов, что свидетельствует о потенциальной длительной активации аденозиновых рецепторов А1.
Кардиология и хирургия
Было показано, что АТФ является безопасным и практичным легочным сосудорасширяющим средством у пациентов с легочной гипертензией. [21] Точно так же аденозин и АТФ можно использовать во время операции, чтобы вызвать гипотензию у пациентов.[21]
[21] Точно так же аденозин и АТФ можно использовать во время операции, чтобы вызвать гипотензию у пациентов.[21]
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Литература
- 1.
Meurer F, Do HT, Sadowski G, Held C. Стандартная энергия Гиббса метаболических реакций: II. Глюкозо-6-фосфатазная реакция и гидролиз АТФ. Биофиз хим. 2017 Апр; 223:30-38. [PubMed: 28282626]
- 2.
Beis I, Newsholme EA. Содержание адениновых нуклеотидов, фосфагенов и некоторых гликолитических интермедиатов в покоящихся мышцах позвоночных и беспозвоночных. Биохим Дж. 1975 октября; 152(1):23-32. [Бесплатная статья PMC: PMC1172435] [PubMed: 1212224]
- 3.
Wang X, Zhang X, Wu D, Huang Z, Hou T, Jian C, Yu P, Lu F, Zhang R, Sun T, Li J, Qi W, Wang Y, Gao F, Cheng H. Митохондриальные вспышки регулируют гомеостаз АТФ в сердце.
 Элиф. 2017 Jul 10;6 [PMC бесплатная статья: PMC5503511] [PubMed: 28692422]
Элиф. 2017 Jul 10;6 [PMC бесплатная статья: PMC5503511] [PubMed: 28692422]- 4.
Mishra NS, Tuteja R, Tuteja N. Передача сигналов через сети киназ MAP в растениях. Арх Биохим Биофиз. 2006 01 августа; 452 (1): 55-68. [В паблике: 16806044]
- 5.
Лин Х, Айрапетов М.К., Сун Г. Характеристика взаимодействий между активным центром протеинтирозинкиназы и активатором двухвалентного металла. БМС Биохим. 2005 23 ноября; 6:25. [PMC бесплатная статья: PMC1316873] [PubMed: 16305747]
- 6.
Zimmermann H. Внеклеточный АТФ и другие нуклеотиды-повсеместные триггеры высвобождения межклеточного мессенджера. Пуринергический сигнал. 2016 март; 12(1):25-57. [Бесплатная статья PMC: PMC4749530] [PubMed: 26545760]
- 7.
Каменецкий М., Миддельхауфе С., Банк Э.М., Левин Л.Р., Бак Дж., Стигборн С. Молекулярные детали образования цАМФ в клетках млекопитающих: рассказ о двух системах. Дж Мол Биол. 2006 г., 29 сентября; 362(4):623-39.
 [Бесплатная статья PMC: PMC3662476] [PubMed: 16934836]
[Бесплатная статья PMC: PMC3662476] [PubMed: 16934836]- 8.
Джойс CM, Steitz TA. Структуры и функции полимеразы: вариации на тему? J Бактериол. 1995 ноябрь; 177 (22): 6321-9. [Бесплатная статья PMC: PMC177480] [PubMed: 7592405]
- 9.
Bonora M, Patergnani S, Rimessi A, De Marchi E, Suski JM, Bononi A, Giorgi C, Marchi S, Missiroli S, Poletti F, Wieckowski MR, Pinton P. Синтез и хранение АТФ. Пуринергический сигнал. 2012 Сентябрь;8(3):343-57. [Бесплатная статья PMC: PMC3360099] [PubMed: 22528680]
- 10.
Карденас С., Миллер Р.А., Смит И., Буй Т., Молго Дж., Мюллер М., Вайс Х., Чунг К.Х., Ян Дж., Паркер И., Томпсон С.Б., Бирнбаум М.Дж., Халлоус К.Р., Фоскетт Дж.К. Существенная регуляция биоэнергетики клетки путем конститутивного переноса Ca2+ рецептора InsP3 в митохондрии. Клетка. 2010 23 июля; 142 (2): 270-83. [Бесплатная статья PMC: PMC2911450] [PubMed: 20655468]
- 11.
Пабло Хуидобро-Торо Дж.
 , Вероника Доносо М. Симпатическая котрансмиссия: скоординированное действие АТФ и норадреналина и их модуляция нейропептидом Y в сосудистых нейроэффекторных соединениях человека. Евр Дж Фармакол. 2004 01 октября; 500 (1-3): 27-35. [PubMed: 15464018]
, Вероника Доносо М. Симпатическая котрансмиссия: скоординированное действие АТФ и норадреналина и их модуляция нейропептидом Y в сосудистых нейроэффекторных соединениях человека. Евр Дж Фармакол. 2004 01 октября; 500 (1-3): 27-35. [PubMed: 15464018]- 12.
Coco S, Calegari F, Pravettoni E, Pozzi D, Taverna E, Rosa P, Matteoli M, Verderio C. Хранение и высвобождение АТФ из астроцитов в культуре. Дж. Биол. Хим. 2003 10 января; 278 (2): 1354-62. [В паблике: 12414798]
- 13.
Аттвелл Д., Лафлин С.Б. Энергетический баланс для передачи сигналов в сером веществе головного мозга. J Cereb Blood Flow Metab. 21 октября 2001 г. (10): 1133-45. [PubMed: 11598490]
- 14.
Harris JJ, Jolivet R, Attwell D. Использование и поставка синаптической энергии. Нейрон. 06 сентября 2012 г .; 75 (5): 762–777. [PubMed: 22958818]
- 15.
Вонг-Райли MT. Цитохромоксидаза: эндогенный метаболический маркер активности нейронов.
 Тренды Нейроси. 1989 марта; 12(3):94-101. [PubMed: 2469224]
Тренды Нейроси. 1989 марта; 12(3):94-101. [PubMed: 2469224]- 16.
Барклай CJ. Энергетика сокращения. сост. физиол. 2015 Апрель;5(2):961-95. [PubMed: 25880520]
- 17.
Богатый PR. Молекулярный механизм дыхательной цепи Кейлина. Биохим Сок Транс. 2003 г.; 31 декабря (часть 6): 1095–105. [PubMed: 14641005]
- 18.
Ronnett GV, Kim EK, Landree LE, Tu Y. Метаболизм жирных кислот как мишень для лечения ожирения. Физиол Поведение. 2005 19 мая;85(1):25-35. [PubMed: 15878185]
- 19.
Бровко Л.Ю., Романова Н.А., Угарова Н.Н. Биолюминесцентный анализ бактериального внутриклеточного АМФ, АДФ и АТФ с использованием коиммобилизованного трехферментного реагента (аденилаткиназа, пируваткиназа и люцифераза светлячка). Анальная биохимия. 1994 01 августа; 220 (2): 410-4. [PubMed: 7978286]
- 20.
Хаяшида М., Фукуда К., Фукунага А. Клиническое применение аденозина и АТФ для обезболивания.