Гипертрофия скелетных мышц 2023. Биомеханика силовых упражнений — обучение на семинаре в Рассказове Колледжа Вейдера
Гипертрофия скелетных мышц 2023. Биомеханика силовых упражнений — обучение на семинаре в Рассказове Колледжа Вейдера
Документы: сертификат
Продолжительность: 1 день
Время обучения: 8 часов
10 500 ₽*11 200 ₽
Используемый на семинаре материал, разработан на основе анализа большого количества современной российской и зарубежной научной литературы (научные статьи, книги, диссертации), а также собственных исследований А.В. Самсоновой и сотрудников кафедры биомеханики. Каждая лекция сопровождается большим иллюстративным материалом, представленным в виде презентаций.
Последовательно раскрываются кинематические и динамические характеристики двигательных действий. Примеры, используемые на семинаре в Рассказове основаны на анализе силовых упражнений.
Программа семинара
- Состав и строение скелетных мышц, мышечных волокон и миофибрилл
- Типы мышечных волокон. Методы оценки композиции мышечных волокон
- Механизмы миофибриллярной и саркоплазматической гипертрофии мышц
- Влияние различных параметров тренировки на гипертрофию мышц
- Саркопения (влияние возраста на скелетные мышцы)
- Основные понятия биомеханики
- Биомеханика опорно-двигательного аппарата человека
- Биомеханика силовых упражнений. (Сравнительный анализ силовых упражнений по воздействию на скелетные мышцы и суставы)
- Биомеханика травм опорно-двигательного аппарата человека при выполнении силовых упражнений
Выдаваемые документы
Сертификат «Гипертрофия скелетных мышц»
Общие сведения
Гипертрофия скелетных мышц (увеличение объема скелетных мышц) рассматривается с позиций ряда медико-биологических дисциплин: анатомии, биомеханики, биохимии, гистологии, спортивной медицины, цитологии, физиологии, а также теории и методики атлетизма.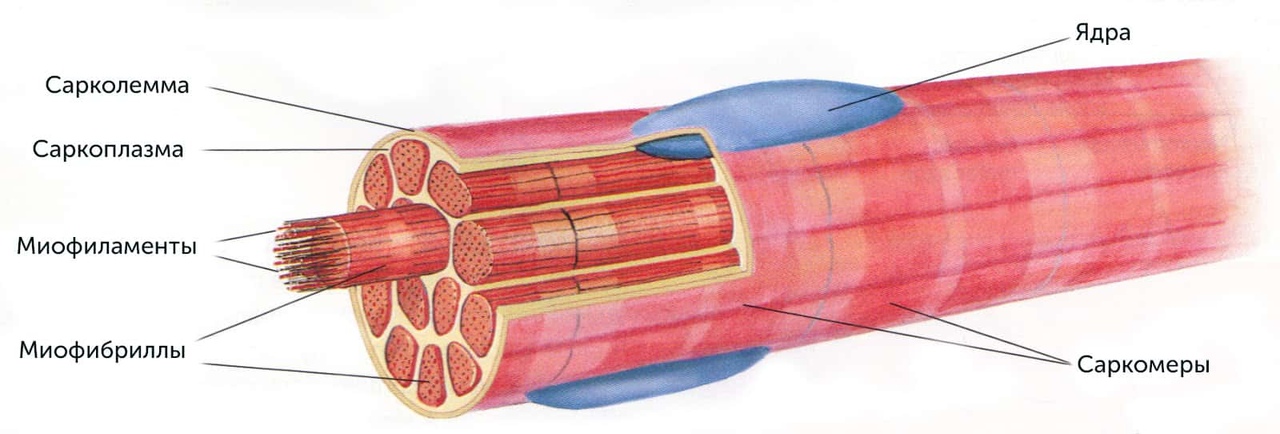 Данные сведения необходимы для понимания биомеханики опорно-двигательного аппарата и факторов, влияющих на развитие силы скелетными мышцами.
Данные сведения необходимы для понимания биомеханики опорно-двигательного аппарата и факторов, влияющих на развитие силы скелетными мышцами.
Информация для поступления и учёбы
При оплате обучения не менее чем за 30 дней до начала действует скидка 5%.
Договор на обучение подписывает один из родителей (опекунов), если на начало обучения, учащемуся менее 18 лет.
Для участия предварительная запись обязательна. Семинар переносится на следующую дату, если не набрано минимальное количество участников.
Регистрация в офисе за час до начала занятий.
Преподаватель
Самсонова Алла Владимировна
Рекомендуем:
Теги: Анатомия, Биомеханика
Задать вопрос
Ждите…
clean
_
Миотония Томсена/Беккера, CLCN1 ч.
 м. — узнать цены на анализ и сдать в Торбеево
м. — узнать цены на анализ и сдать в ТорбеевоМетод определения ПЦР, секвенирование
Исследуемый материал Цельная кровь (с ЭДТА)
Исследование частых мутаций в гене CLCN1.
Тип наследования.
Аутосомно-доминантный, аутосомно-рецессивный.
Гены, ответственные за развитие заболевания.
CLСN1 (CHLORIDE CHANNEL 1, SKELETAL MUSCLE) – ген белка мышечных хлорных каналов, расположен на хромосоме 7 в регионе 7q35, содержит 23 экзона.
Определение заболевания.
Миотония — нервно-мышечное заболевание, характеризующееся наличием мышечной гипертрофии и миотонического феномена — замедленной релаксацией мышцы после ее сокращения (сократившаяся мышца долгое время не расслабляется и затем расслабление происходит крайне медленно).
Патогенез и клиническая картина.
Заболевание встречается в двух вариантах — миотонии Томсена (OMIM: 160800) с аутосомно-доминантным типом наследования и миотонии Беккера (OMIM: 255700) с аутосомно-рецессивным типом наследования. Показано, что заболевания являются аллельными вариантами и эффект доминантности и рецессивности обусловлен различными мутациями в одном и том же гене. Миотония Беккера характеризуются ранней манифестацией и более тяжёлыми клиническими проявлениями Белок мышечных хлорных каналов регулирует электрическую возбудимость мембраны скелетных мышц. В отсутствии и при функциональной неактивности белка нарушается проникновение хлора в мышечное волокно, что приводит к возникновению электрической нестабильности мембраны.
Показано, что заболевания являются аллельными вариантами и эффект доминантности и рецессивности обусловлен различными мутациями в одном и том же гене. Миотония Беккера характеризуются ранней манифестацией и более тяжёлыми клиническими проявлениями Белок мышечных хлорных каналов регулирует электрическую возбудимость мембраны скелетных мышц. В отсутствии и при функциональной неактивности белка нарушается проникновение хлора в мышечное волокно, что приводит к возникновению электрической нестабильности мембраны.
1 вариант — миотония Томсена
Первые симптомы заболевания можно отметить с рождения или в периоде новорожденности. Основным проявлением заболевания является миотонический спазм в различных группах мышц, возникающий после их интенсивного произвольного сокращения. Выраженность спазма наибольшая в начале движения и уменьшается во время повторных мышечных сокращений. Усиление миотонического феномена возникает на холоде, уменьшение — в тепле, во время отдыха и при приеме небольших доз алкоголя.
2 вариант — миотония Беккера
В большинстве случаев заболевание возникает в возрасте от 4 до 12 лет у девочек и около 18 лет у мальчиков. Первые признаки миотонии возникают в мышцах ног, через несколько лет — в мышцах рук.
На электромиограмме выявляется специфический миотонический феномен.
У части больных может быть нерезко выраженное увеличение уровня активности креатинфосфокиназы в плазме крови.
Специфические морфологические признаки отсутствуют. В большинстве случаев выявляется вариабельный диаметр мышечных волокон, их гипертрофия и централизация ядер.
Частота встречаемости: в Европе оценивается как 1:100 000 семей, в странах Скандинавии – 1:10 000.
Перечень исследуемых мутаций может быть предоставлен по запросу.
Прием фосфатидной кислоты увеличивает гипертрофию и силу скелетных мышц | Журнал Международного общества спортивного питания
Добавка фосфатидной кислоты увеличивает гипертрофию и силу скелетных мышц
Скачать PDF
Скачать ePub
Том 10, Приложение 1
- Постерная презентация
- Открытый доступ
- Опубликовано:
- Джордан М Джой 1 ,
- Райан П Лоури 1 ,
- Джошуа Э Дудек 1 ,
- Эдуардо О Де Соуза 2 ,
- Ральф Ягер 3 ,
- Шон А Макклири 1 ,
- Стефани М. К. Уилсон 4 ,
- Мартин Пурпура 3 и
- …
- Джейкоб М. Уилсон 1
Журнал Международного общества спортивного питания том 10 , Номер статьи: P13 (2013) Процитировать эту статью
15 тыс. обращений
15 Альтметрический
Введение
Срастание ткани скелетных мышц может иметь решающее значение для различных групп населения, включая спортсменов и пожилых людей. Гипертрофия скелетных мышц в значительной степени опосредована усилением синтеза мышечного белка. Было показано, что мишень рапамицина у млекопитающих (mTOR) регулирует скорость синтеза мышечного белка, а механический стимул (упражнения с отягощениями) активирует mTOR, ключевую роль в котором играет фосфолипидная фосфатидная кислота (PA). Первое пилотное исследование показало, что пероральный прием ПА, полученного из сои, у спортсменов, проходящих прогрессивную тренировку с отягощениями, скорее всего, привел к большему увеличению силы приседаний и мышечной массы по сравнению с плацебо. Тем не менее, это пилотное исследование, вероятно, было недостаточно мощным, тренировки не проводились под наблюдением и не проводились прямые измерения гипертрофии скелетных мышц. Таким образом, целью данного исследования было изучение влияния ФА на состав тела, силу, мощь и мышечную гипертрофию.
Гипертрофия скелетных мышц в значительной степени опосредована усилением синтеза мышечного белка. Было показано, что мишень рапамицина у млекопитающих (mTOR) регулирует скорость синтеза мышечного белка, а механический стимул (упражнения с отягощениями) активирует mTOR, ключевую роль в котором играет фосфолипидная фосфатидная кислота (PA). Первое пилотное исследование показало, что пероральный прием ПА, полученного из сои, у спортсменов, проходящих прогрессивную тренировку с отягощениями, скорее всего, привел к большему увеличению силы приседаний и мышечной массы по сравнению с плацебо. Тем не менее, это пилотное исследование, вероятно, было недостаточно мощным, тренировки не проводились под наблюдением и не проводились прямые измерения гипертрофии скелетных мышц. Таким образом, целью данного исследования было изучение влияния ФА на состав тела, силу, мощь и мышечную гипертрофию.
Методы
В этом исследовании приняли участие 28 мужчин, тренирующихся с отягощениями (возраст 21 ± 3 года, масса тела 76 ± 9 кг и рост 176 см ± 9 см). Субъекты были поровну разделены на экспериментальные и контрольные условия, и каждый субъект принимал участие в 8-недельной периодической программе тренировок с отягощениями. Программа тренировок с отягощениями состояла из двух тренировок в неделю, ориентированных на гипертрофию, и одной тренировки, ориентированной на силу, в неделю. Экспериментальные условия (EXP) получали 750 мг PA, полученного из сои (Mediator™, Chemi Nutra, White Bear Lake, MN), в то время как контрольные условия (CON) получали визуально идентичное плацебо (рисовая мука). Измерения определяемого DEXA состава тела, площади поперечного сечения прямой мышцы бедра, силы 1ПМ и анаэробной мощности проводились до и после 8-недельного тренировочного вмешательства. Повторные измерения 2×2 ANOVA использовали для определения группового, временного и группового х временного взаимодействий. Для обнаружения различий использовали апостериорный анализ Тьюки.
Субъекты были поровну разделены на экспериментальные и контрольные условия, и каждый субъект принимал участие в 8-недельной периодической программе тренировок с отягощениями. Программа тренировок с отягощениями состояла из двух тренировок в неделю, ориентированных на гипертрофию, и одной тренировки, ориентированной на силу, в неделю. Экспериментальные условия (EXP) получали 750 мг PA, полученного из сои (Mediator™, Chemi Nutra, White Bear Lake, MN), в то время как контрольные условия (CON) получали визуально идентичное плацебо (рисовая мука). Измерения определяемого DEXA состава тела, площади поперечного сечения прямой мышцы бедра, силы 1ПМ и анаэробной мощности проводились до и после 8-недельного тренировочного вмешательства. Повторные измерения 2×2 ANOVA использовали для определения группового, временного и группового х временного взаимодействий. Для обнаружения различий использовали апостериорный анализ Тьюки.
Результаты
Наблюдался значительный эффект группы x времени (p=0,02) для CSA, в котором группа EXP увеличилась (+1,01 см 2 , ES = 0,92) в большей степени, чем группа CON (+0,61). см 2 , ES = 0,52). Был значительный групповой эффект времени (p = 0,01) для LBM, в котором группа EXP (+2,4 кг, ES = 0,42) удвоила эффект от одной тренировки с отягощениями (CON +1,2 кг, ES = 0,26). Был значительный эффект группы x времени (p = 0,04) для жима ногами 1RM, в котором группа EXP увеличилась в большей степени (+52,0 кг, ES = 1,2), чем группа CON (+32,5 кг, ES = 0,78). . Для потери жира наблюдался эффект «группа тренда х время» (p = 0,06), в котором группа EXP уменьшила жировые отложения в большей степени, чем группа CON (-1,3 кг против -0,5 кг).
см 2 , ES = 0,52). Был значительный групповой эффект времени (p = 0,01) для LBM, в котором группа EXP (+2,4 кг, ES = 0,42) удвоила эффект от одной тренировки с отягощениями (CON +1,2 кг, ES = 0,26). Был значительный эффект группы x времени (p = 0,04) для жима ногами 1RM, в котором группа EXP увеличилась в большей степени (+52,0 кг, ES = 1,2), чем группа CON (+32,5 кг, ES = 0,78). . Для потери жира наблюдался эффект «группа тренда х время» (p = 0,06), в котором группа EXP уменьшила жировые отложения в большей степени, чем группа CON (-1,3 кг против -0,5 кг).
Заключение
Добавка с PA, полученным из сои, может улучшить реакцию гипертрофии скелетных мышц, безжировую массу тела и максимальную силу.
Благодарности
При поддержке Chemi Nutra, White Bear Lake, Миннесота, США.
Информация об авторе
Авторы и организации
Департамент медицинских наук и деятельности человека, Университет Тампы, Тампа, Флорида, США
Jordan M Joy, Ryan P Lowery, Joshua E Dudeck, Sean A McCleary и Джейкоб М Уилсон
Лаборатория нейромышечной адаптации к силовым тренировкам, Школа физического воспитания и спорта, Университет Сан-Паулу, Сан-Паулу, Бразилия
Eduardo O De Souza
Increnovo LLC, 2138 E Lafayette P л, Милуоки, Висконсин, США
Ральф Ягер и Мартин Пурпура
Департамент питания, Институт производительности IMG, Академия IMG, Брадентон, Флорида, США
Стефани М.
 С. Уилсон
С. Уилсон
- Jordan M Joy
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ryan P Lowery
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Joshua E Dudeck
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Eduardo O De Souza
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ralf Jäger
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Sean A McCleary
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Стефани М. С. Уилсон
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Martin Purpura
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Jacob M Wilson
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за переписку
Связь с
Джейкоб М Уилсон.
Права и разрешения
Открытый доступ Эта статья опубликована по лицензии компании BioMed Central Ltd. Эта статья находится в открытом доступе и распространяется в соответствии с условиями лицензии Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), который разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии надлежащего указания оригинальной работы. Отказ от права Creative Commons на общественное достояние ( https://creativecommons.org/publicdomain/zero/1.0/ ) применяется к данным, представленным в этой статье, если не указано иное.
Перепечатки и разрешения
Об этой статье
Понимание гипертрофии скелетных мышц: интеграция клеточных сигналов
Понимание гипертрофии скелетных мышц: интеграция клеточных сигналов
Все физиологи знают, что мышечная активность необходима для поддержания мышечной массы.
 Но какие основные молекулярные механизмы работают в мышцах? Здесь Дуглас Болстер и его коллеги предлагают некоторые идеи 90 107
Но какие основные молекулярные механизмы работают в мышцах? Здесь Дуглас Болстер и его коллеги предлагают некоторые идеи 90 107Характеристики
Дуглас Р. Болстер (1), Нил Кубица (1), Стивен Дж. Крозье (1), Дэвид Л. Уильямсон (1), Питер А. Фаррелл (2), Скот Р. Кимбалл (1) и Леонард С. Джефферсон (1)
1: Медицинский колледж Пенсильванского государственного университета, кафедра клеточной и молекулярной физиологии, Херши, Пенсильвания, США
2: Университет Восточной Каролины, кафедра физических упражнений и спортивных наук, Гринвилл, Северная Каролина, США
https://doi.org/10.36866/pn.55.18
первый ряд (слева направо): Леонард Джефферсон и Дуглас Болстер; задний ряд (слева направо): Дэвид Уильямсон, Стивен Крозье, Нил Кубица и Скот Кимбалл.
Peter Farrell
Жизненно важное значение скелетных мышц для общего состояния здоровья и повседневной деятельности, вероятно, воспринимается большинством людей как должное. Скелетные мышцы необходимы для базовой осанки, движения и различных метаболических функций. В частности, скелетные мышцы составляют примерно 40-50% от общей массы тела, служат преобладающим местом метаболизма глюкозы и в значительной степени способствуют основному метаболизму. Поддержание или даже увеличение массы скелетных мышц становится критически важным в контексте старения (саркопения) и различных болезненных состояний, связанных с потерей мышечной массы (например, сепсис, рак, диабет, ВИЧ). Таким образом, в последнее время были предприняты значительные усилия для выяснения клеточных и молекулярных механизмов, посредством которых происходит потеря (атрофия) и увеличение (гипертрофия) скелетных мышц. Важно отметить, что процессы, вызывающие мышечную атрофию, относительно уникальны и не являются простой противоположностью гипертрофии.
Скелетные мышцы необходимы для базовой осанки, движения и различных метаболических функций. В частности, скелетные мышцы составляют примерно 40-50% от общей массы тела, служат преобладающим местом метаболизма глюкозы и в значительной степени способствуют основному метаболизму. Поддержание или даже увеличение массы скелетных мышц становится критически важным в контексте старения (саркопения) и различных болезненных состояний, связанных с потерей мышечной массы (например, сепсис, рак, диабет, ВИЧ). Таким образом, в последнее время были предприняты значительные усилия для выяснения клеточных и молекулярных механизмов, посредством которых происходит потеря (атрофия) и увеличение (гипертрофия) скелетных мышц. Важно отметить, что процессы, вызывающие мышечную атрофию, относительно уникальны и не являются простой противоположностью гипертрофии.
Увеличение массы скелетных мышц обусловлено процессом обмена белка, который представляет собой баланс между синтезом белка и его распадом. Для достижения гипертрофии необходимо поддерживать более высокие скорости синтеза белка по сравнению с расщеплением белка, в то время как повышенный распад белка вызывает потерю белка. В целом, эти процессы происходят непрерывно и подвержены внешней модуляции такими факторами, как доступность питательных веществ, гормоны и физические упражнения.
В целом, эти процессы происходят непрерывно и подвержены внешней модуляции такими факторами, как доступность питательных веществ, гормоны и физические упражнения.
Молекулярная регуляция роста
Молекулярный контроль, который управляет изменениями в синтезе белка и возможным увеличением мышечной массы, включает как транскрипционные, так и трансляционные входы. Хотя бывают исключения, последствия, связанные с измененной транскрипцией гена, обычно проявляются в течение периода от нескольких дней до недель, тогда как эффекты, связанные с трансляцией мРНК (т.е. процесс синтеза белка на основе информации, закодированной мРНК), могут проявляться в течение нескольких минут или часов. . Каждая из транскрипций и трансляций состоит из трех отдельных стадий (инициация, элонгация, терминация) с преобладающей регуляцией в фазе инициации. Однако трансляция уникальна, потому что мРНК рекрутируется, а не продуцируется, и этот процесс реагирует на острые метаболические/пищевые изменения. В центре внимания настоящей статьи будет освещение влияния инициации трансляции мРНК на острые изменения в синтезе белка, активация отдельных мРНК, связанных с ростом, и то, как эти события приводят к изменению экспрессии генов в контексте упражнений с отягощениями.
В центре внимания настоящей статьи будет освещение влияния инициации трансляции мРНК на острые изменения в синтезе белка, активация отдельных мРНК, связанных с ростом, и то, как эти события приводят к изменению экспрессии генов в контексте упражнений с отягощениями.
Инициация трансляции и упражнения с сопротивлением
Инициация трансляции по существу включает два центральных компонента, опосредованных эукариотическими факторами инициации (eIF), которые контролируют события, ограничивающие скорость. Эти два компонента, говоря простым языком, позволяют рибосоме связываться с мРНК (комплекс eIF4F) и доставлять рибосому к месту на мРНК, где начинается трансляция (eIF2/eIF2B) (рис. 1). Существенный механизм регуляции роста при инициации трансляции включает белок «мишень рапамицина» (mTOR) млекопитающих. Двумя распространенными нижестоящими мишенями mTOR являются 70-кДа рибосомальная протеинкиназа S6 (S6K1) и eIF4E-связывающий белок-1 (4E-BP1).
Рисунок 1. Поступательный контроль синтеза белков скелетных мышц после упражнений с отягощениями. Передача сигналов через eIF2/eIF2B, по-видимому, контролирует резкое увеличение общей скорости синтеза белка после упражнений с отягощениями. Как путь mTOR может регулировать eIF2B, в настоящее время неизвестно. Активация белков S6K1 и eIF4F в основном отвечает за увеличение способности синтезировать белок при хронических тренировках с отягощениями.
Передача сигналов через eIF2/eIF2B, по-видимому, контролирует резкое увеличение общей скорости синтеза белка после упражнений с отягощениями. Как путь mTOR может регулировать eIF2B, в настоящее время неизвестно. Активация белков S6K1 и eIF4F в основном отвечает за увеличение способности синтезировать белок при хронических тренировках с отягощениями.
Распространенное заблуждение относительно изменений в инициации трансляции заключается в том, что активация любого белка в этом пути соответствует увеличению синтеза белка. Например, после упражнений с отягощениями повышение синтеза белка задерживается на несколько часов, в то время как mTOR-опосредованные события могут быстро усиливаться в течение этого периода (Nader & Esser, 2001). Возможное увеличение синтеза белка, по-видимому, совпадает с более поздними изменениями eIF2B (Farrell и др. . 1999). Без сомнения, хроническая передача сигналов mTOR необходима для обеспечения увеличения размера клеток/мышечной массы, поскольку ингибирование этого пути почти полностью блокирует ответ (Bodine et al , 2001). Кроме того, нижняя мишень mTOR, S6K1, тесно связана с мышечной гипертрофией (Baar & Esser, 1999). Однако резкое подавление mTOR, по-видимому, не оказывает существенного влияния на общую скорость синтеза белка в скелетных мышцах. В совокупности правильно предположить, что оба компонента инициации трансляции необходимы для увеличения массы скелетных мышц. События, связанные с регуляцией eIF2B, могут управлять резкими изменениями в синтезе белка после упражнений с отягощениями, в то время как активация mTOR/4E-BP1/S6K1 может приводить к преимущественному синтезу белков, необходимых для усиления аппарата трансляции и оптимизации способности к синтезу белка при длительных нагрузках. срок обучения.
Кроме того, нижняя мишень mTOR, S6K1, тесно связана с мышечной гипертрофией (Baar & Esser, 1999). Однако резкое подавление mTOR, по-видимому, не оказывает существенного влияния на общую скорость синтеза белка в скелетных мышцах. В совокупности правильно предположить, что оба компонента инициации трансляции необходимы для увеличения массы скелетных мышц. События, связанные с регуляцией eIF2B, могут управлять резкими изменениями в синтезе белка после упражнений с отягощениями, в то время как активация mTOR/4E-BP1/S6K1 может приводить к преимущественному синтезу белков, необходимых для усиления аппарата трансляции и оптимизации способности к синтезу белка при длительных нагрузках. срок обучения.
Быстрая передача клеточных сигналов и упражнения с отягощениями
Недавние попытки лучше понять регуляцию инициации трансляции после острого приступа упражнений с отягощениями предполагают изменения, при которых отдельные белки eIF быстро фосфорилируются (Bolster et al. 2003). Прерывистая и временная активация этих белков может обеспечить более точный контроль модуляции реакции роста. В частности, эти ответы кажутся временными по своей природе, и острое воздействие упражнений с отягощениями на трансляцию мРНК, вероятно, становится кумулятивным с каждым последующим циклом упражнений; подразумевается, что этот путь роста периодически «включается» при повторяющихся упражнениях с отягощениями, и различные мРНК (рибосомные белки и т. д.) могут накапливаться до такой степени, что происходит увеличение количества специфических белков (Neufer & Dohm, 19).93). Эти ответы подчеркивают долгосрочные и более быстрые механизмы контроля, связанные с транскрипцией и трансляцией, соответственно, которые способствуют достижению мышечной гипертрофии (Fig. 2).
Прерывистая и временная активация этих белков может обеспечить более точный контроль модуляции реакции роста. В частности, эти ответы кажутся временными по своей природе, и острое воздействие упражнений с отягощениями на трансляцию мРНК, вероятно, становится кумулятивным с каждым последующим циклом упражнений; подразумевается, что этот путь роста периодически «включается» при повторяющихся упражнениях с отягощениями, и различные мРНК (рибосомные белки и т. д.) могут накапливаться до такой степени, что происходит увеличение количества специфических белков (Neufer & Dohm, 19).93). Эти ответы подчеркивают долгосрочные и более быстрые механизмы контроля, связанные с транскрипцией и трансляцией, соответственно, которые способствуют достижению мышечной гипертрофии (Fig. 2).
Рисунок 2. Предлагаемая модель клеточной адаптации при упражнениях с отягощениями. Непосредственный период восстановления после упражнений с отягощениями включает трансляционную регуляцию, посредством которой активируются отдельные белки eIF и активируются специфические связанные с ростом мРНК. Повторяющиеся упражнения с отягощениями включают регуляцию транскрипции, теоретически накапливаются специфические мРНК и синтезируются новые белки. Трансляция и транскрипция мРНК работают вместе, чтобы координировать возможное увеличение массы скелетных мышц. (E: упражнение; неактивная мРНК: становится активной после каждого последующего упражнения).
Повторяющиеся упражнения с отягощениями включают регуляцию транскрипции, теоретически накапливаются специфические мРНК и синтезируются новые белки. Трансляция и транскрипция мРНК работают вместе, чтобы координировать возможное увеличение массы скелетных мышц. (E: упражнение; неактивная мРНК: становится активной после каждого последующего упражнения).
Куда нам двигаться дальше?
Идентификация ключевых факторов, которые быстро инициируют каскад сигнальных событий в ответ на острые упражнения с отягощениями, остается неуловимой, но некоторые кандидаты могут включать активацию интегрина и/или мобилизацию кальция. Интегрины представляют собой трансмембранные белки, которые связывают физические или химические стимулы с внутриклеточными событиями, тогда как повышенный поток кальция проявляется во время мышечного сокращения. Быстрая, но временная активация пути, продемонстрированная упражнениями с отягощениями, предполагает, что эти факторы могут быстро стимулировать передачу сигналов mTOR, что затем может привести к селективной трансляции мРНК локальных факторов роста (т.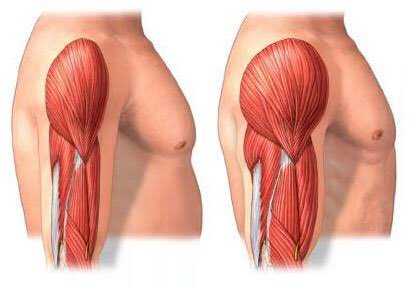 е. IGF-1), дополнительно усиливая реакцию роста клетки.
е. IGF-1), дополнительно усиливая реакцию роста клетки.
Поскольку мы смотрим в будущее, чтобы понять, как будет развиваться наше понимание мышечной гипертрофии, важно также осознавать проблемы. Сравнение данных в этой области исследований часто затруднено из-за множества используемых в настоящее время моделей гипертрофии. Не всегда существует тесная корреляция между культурой клеток и данными животных или человека. Более того, перекрытие различных сигнальных путей и быстро расширяющееся участие вновь идентифицированных белков усложняют интерпретацию. Реакция клетки на рост включает несколько входных сигналов. Поэтому требуется интегрированный ответ, и, как бы заманчиво это ни было, гипертрофия, скорее всего, никогда не будет изолирована от одного ключевого белка, действующего как «главный переключатель». Таким образом, многогранный подход с использованием геномных, протеомных и биоинформационных инструментов будет необходим для выяснения пробелов в наших знаниях о том, как происходит мышечная гипертрофия.
Ссылки
Баар К. и Эссер К.А. (1999). Фосфорилирование p70S6k коррелирует с увеличением массы скелетных мышц после упражнений с отягощениями. Am J Physiol 276 , C120-C127.
Бодин С.К., Ститт Т.Н., Гонсалес М., Клайн В.О., Стовер Г.Л., Бауэрлейн Р., Злотченко Э., Скримджер А., Лоуренс Дж.К., Гласс Д.Дж. и Янкопулус Г.Д. (2001). Путь Akt/mTOR является важным регулятором гипертрофии скелетных мышц и может предотвращать мышечную атрофию 90–250 in vivo . Nature Cell Biol 3 , 1014-1019.
Болстер Д.Р., Кубица Н., Крозье С.Дж., Уильямсон Д.Л., Фаррелл П.А., Кимбалл С.Р. и Джефферсон Л.С. (2003). Немедленный ответ мишени рапамицина (mTOR) у млекопитающих, опосредованной передачей сигналов после острых упражнений с отягощениями в скелетных мышцах крыс. Дж Физиол 553 .1, 213-220.
Farrell PA, Fedele MJ, Vary TC, Kimball SR, Lang CH & Jefferson LS (1999). Регуляция синтеза белка после острых упражнений с отягощениями у крыс с диабетом.
