— источники энергии — Биохимия
Способы получения энергии в клетке
В клетке существуют четыре основных процесса, обеспечивающих высвобождение энергии из химических связей при окислении веществ и ее запасание:
1. Гликолиз (2 этап биологического окисления) – окисление молекулы глюкозы до двух молекул пировиноградной кислоты, при этом образуется 2 молекулы АТФ и НАДН. Далее пировиноградная кислота в аэробных условиях превращается в ацетил-SКоА, в анаэробных условиях – в молочную кислоту.
2. β-Окисление жирных кислот (2 этап биологического окисления) – окисление жирных кислот до ацетил-SКоА, здесь образуются молекулы НАДН и ФАДН2. Молекулы АТФ «в чистом виде» не появляются.
3. Цикл трикарбоновых кислот (ЦТК, 3 этап биологического окисления) – окисление ацетильной группы (в составе ацетил-SКоА) или иных кетокислот до углекислого газа. Реакции полного цикла сопровождаются образованием 1 молекулы ГТФ (что эквивалентно одной АТФ), 3 молекул 
4. Окислительное фосфорилирование (3 этап биологического окисления) – окисляются НАДН и ФАДН2, полученные в реакциях катаболизма глюкозы, аминокислот и жирных кислот. При этом ферменты дыхательной цепи на внутренней мембране митохондрий обеспечивают образование большей части клеточного АТФ.
Два способа синтеза АТФ
В клетке постоянно происходит использование всех нуклеозидтрифосфатов (АТФ, ГТФ, ЦТФ, УТФ, ТТФ) как донора энергии. При этом АТФ является универсальным макроэргом, участвующим практически во всех сторонах метаболизма и деятельности клетки. И именно за счет АТФ обеспечивается фосфорилирование нуклеотидов ГМФ и ГДФ, ЦДФ, УМФ и УДФ, ТМФ и ТДФ до нуклеозидтрифосфатов.
1. Основным способом получения АТФ в клетке является окислительное фосфорилирование, протекающее в структурах внутренней мембраны митохондрий. При этом энергия атомов водорода молекул НАДН и ФАДН 2, образованных в гликолизе и ЦТК, при окислении жирных кислот и аминокислот, преобразуется в энергию связей АТФ.
2. Однако также есть другой способ фосфорилирования АДФ до АТФ – субстратное фосфорилирование. Этот способ связан с передачей макроэргического фосфата или энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся метаболиты гликолиза (1,3-дифосфоглицериновая кислота, фосфоенолпируват), цикла трикарбоновых кислот (сукцинил-SКоА) и резервный макроэрг креатинфосфат. Энергия гидролиза их макроэргической связи выше, чем 7,3 ккал/моль в АТФ, и роль указанных веществ сводится к использованию этой энергии для фосфорилирования молекулы АДФ до АТФ.
Классификация макроэргов
Макроэргические соединения классифицируются по типу связи, несущей дополнительную энергию:
1. Фосфоангидридная связь. Такую связь имеют все нуклеотиды: нуклеозидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ, ТТФ) и нуклеозиддифосфаты (АДФ, ГДФ, ЦДФ, УДФ, ТДФ).
2. Тиоэфирная связь.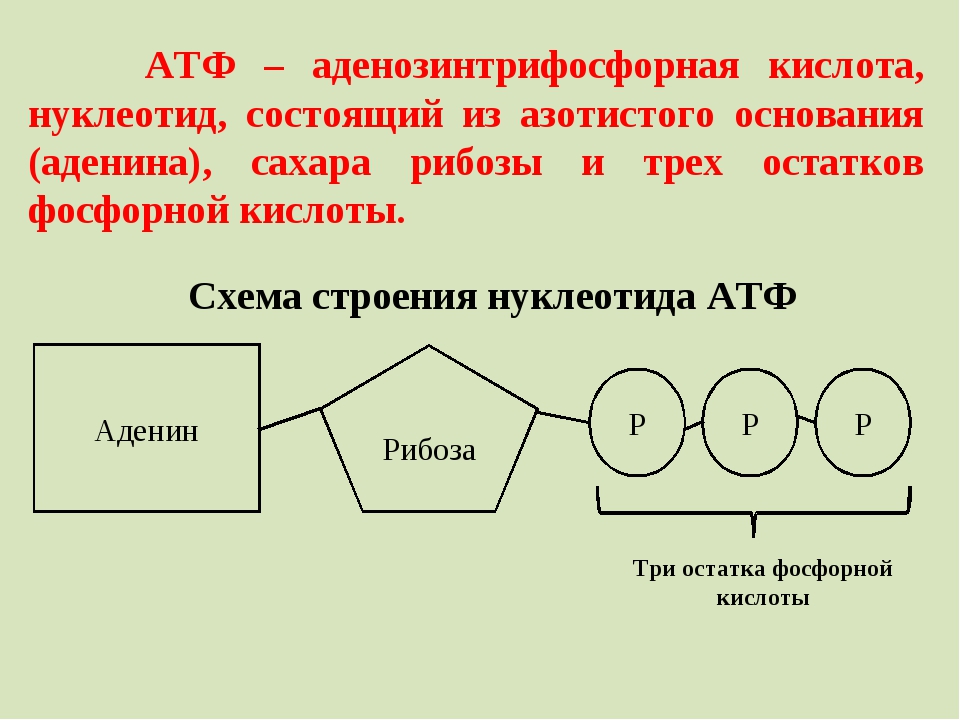 Примером являются ацил-производные коэнзима А: ацетил-SКоА, сукцинил-SКоА, и другие соединения любой жирной кислоты c HS-КоА.
Примером являются ацил-производные коэнзима А: ацетил-SКоА, сукцинил-SКоА, и другие соединения любой жирной кислоты c HS-КоА.
3. Гуанидинфосфатная связь – присутствует в креатинфосфате, запасном макроэрге мышечной и нервной ткани.
4. Ацилфосфатная связь. К таким макроэргам относится метаболит гликолиза 1,3-дифосфоглицериновая кислота (1,3-дифосфоглицерат). Она обеспечивает синтез АТФ в реакции субстратного фосфорилирования.
5. Енолфосфатная
связь. Представитель – фосфоенолпируват, метаболит гликолиза. Он также обеспечивает синтез АТФ в реакции субстратного фосфорилирования в гликолизе.
АТФ-синтаза — ATP synthase — qaz.wiki
Фермент
АТФ-синтаза — это фермент, который катализирует образование молекулы-накопителя энергии аденозинтрифосфата (АТФ) с использованием аденозиндифосфата (АДФ) и неорганического фосфата (P i ). Общая реакция, катализируемая АТФ-синтазой:
- АДФ + Р я + 3H + из ⇌ АТФ + Н 2 О + 3H + в
Образование АТФ из АДФ и P i энергетически невыгодно и обычно протекает в обратном направлении.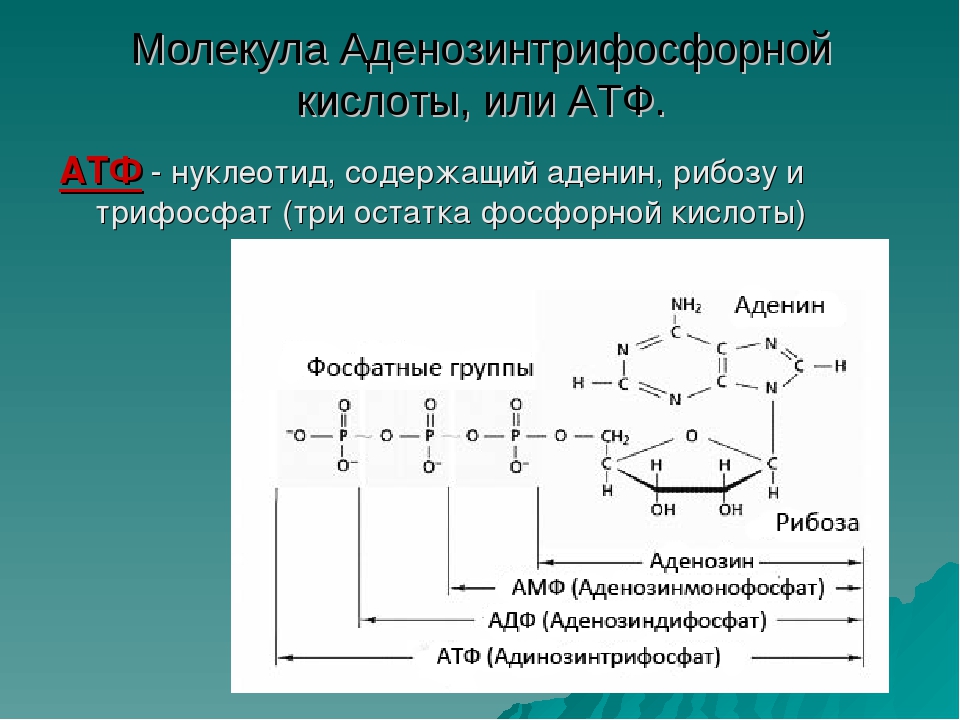 Чтобы ускорить эту реакцию, АТФ-синтаза связывает синтез АТФ во время клеточного дыхания с электрохимическим градиентом, создаваемым разницей в концентрации протонов (H + ) на внутренней митохондриальной мембране у эукариот или плазматической мембране у бактерий. Во время фотосинтеза у растений АТФ синтезируется АТФ-синтазой с использованием протонного градиента, создаваемого в просвете тилакоидов через тилакоидную мембрану и в строму хлоропластов .
Чтобы ускорить эту реакцию, АТФ-синтаза связывает синтез АТФ во время клеточного дыхания с электрохимическим градиентом, создаваемым разницей в концентрации протонов (H + ) на внутренней митохондриальной мембране у эукариот или плазматической мембране у бактерий. Во время фотосинтеза у растений АТФ синтезируется АТФ-синтазой с использованием протонного градиента, создаваемого в просвете тилакоидов через тилакоидную мембрану и в строму хлоропластов .
Эукариотические АТФ-синтазы — это F-АТФазы , работающие «наоборот» для АТФазы . Данная статья посвящена в основном этому типу. F-АТФаза состоит из двух основных субъединиц, F O и F 1 , которые имеют вращательный моторный механизм, позволяющий производить АТФ. Из-за своей вращающейся субъединицы АТФ-синтаза представляет собой молекулярную машину .
Номенклатура
Фракция F 1 получила свое название от термина «Фракция 1», а F O (записанная как нижняя буква «o», а не «ноль») получила свое название от фракции связывания олигомицина , типа природного антибиотика, который способен ингибировать F O- звено АТФ-синтазы.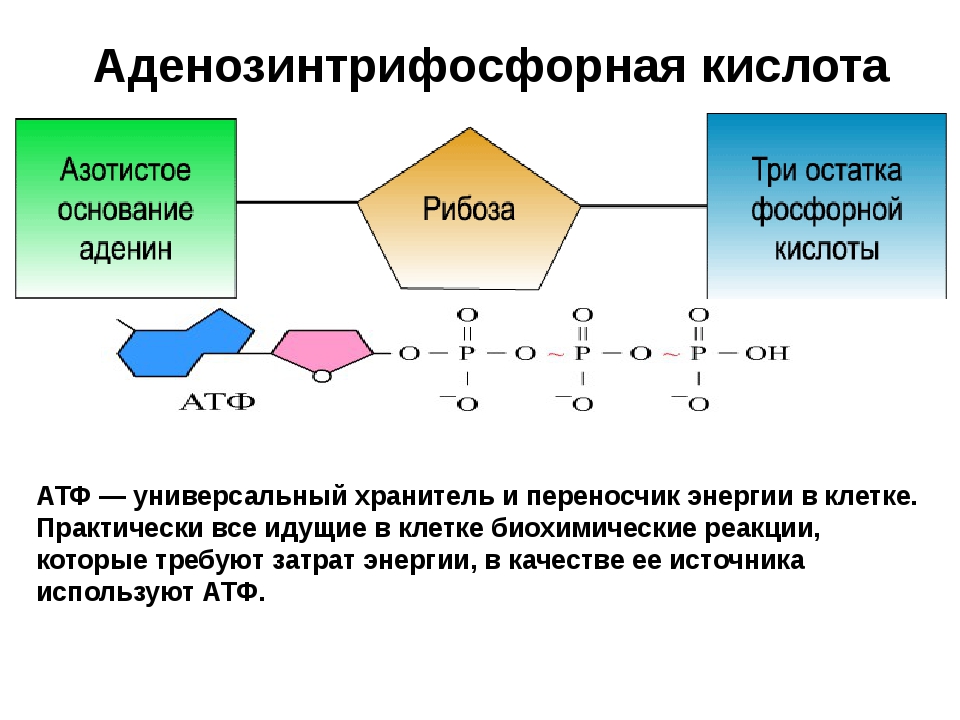
Структура и функции
Митохондриальная АТФ-синтаза крупного рогатого скота. Области F O , F 1 , оси и статора имеют цветовую кодировку пурпурного, зеленого, оранжевого и голубого цветов соответственно. Упрощенная модель F O F 1 -АТФазы, называемой АТФ-синтазой E. coli . Соответственно помечены субъединицы фермента. Двигатель вращения АТФ-синтазы.Расположенная внутри тилакоидной мембраны и внутренней митохондриальной мембраны , АТФ-синтаза состоит из двух областей F O и F 1 . F O вызывает вращение F 1 и состоит из c-кольца и субъединиц a, two b, F6. F 1 состоит из субъединиц α, β, γ, δ. F 1 имеет водорастворимую часть, которая может гидролизовать АТФ. F O, с другой стороны, имеет в основном гидрофобные области.
F 1 регион
Часть F 1 АТФ-синтазы является гидрофильной и отвечает за гидролиз АТФ. Единица F 1 выступает в пространство митохондриального матрикса. Субъединицы α и β образуют гексамер с 6 сайтами связывания. Три из них каталитически неактивны и связывают АДФ.
Остальные три субъединицы катализируют синтез АТФ. Остальные субъединицы F 1 γ, δ, ε являются частью механизма вращения двигателя (ротор / ось). Субъединица γ позволяет β проходить через конформационные изменения (т.е. закрытое, полуоткрытое и открытое состояния), которые позволяют АТФ связываться и высвобождаться после синтеза. Частица F 1 крупная, и ее можно увидеть в просвечивающем электронном микроскопе при отрицательном окрашивании. Это частицы диаметром 9 нм, пронизывающие внутреннюю митохондриальную мембрану.
F O регион
F O субъединица F6 из периферического стебля АТФ-синтазы.
F O представляет собой нерастворимый в воде белок с восемью субъединицами и трансмембранным кольцом. Кольцо имеет форму тетрамера с белком спиральной петли, который претерпевает конформационные изменения при протонировании и депротонировании, подталкивая соседние субъединицы к вращению, вызывая вращение F O, которое затем также влияет на конформацию F 1 , что приводит к переключению состояний альфа и бета-субъединицы. Область FO АТФ-синтазы представляет собой протонную пору, встроенную в митохондриальную мембрану. Он состоит из трех основных субъединиц: a, b и c. Шесть субъединиц c образуют кольцо ротора, а субъединица b образует стержень, соединяющийся с F 1 OSCP, который предотвращает вращение гексамера αβ. Субъединица a соединяет b с кольцом c. У людей есть шесть дополнительных субъединиц: d , e , f , g , F6 и 8 (или A6L). Эта часть фермента расположена на внутренней мембране митохондрий и связывает транслокацию протонов с вращением, которое вызывает синтез АТФ в области F 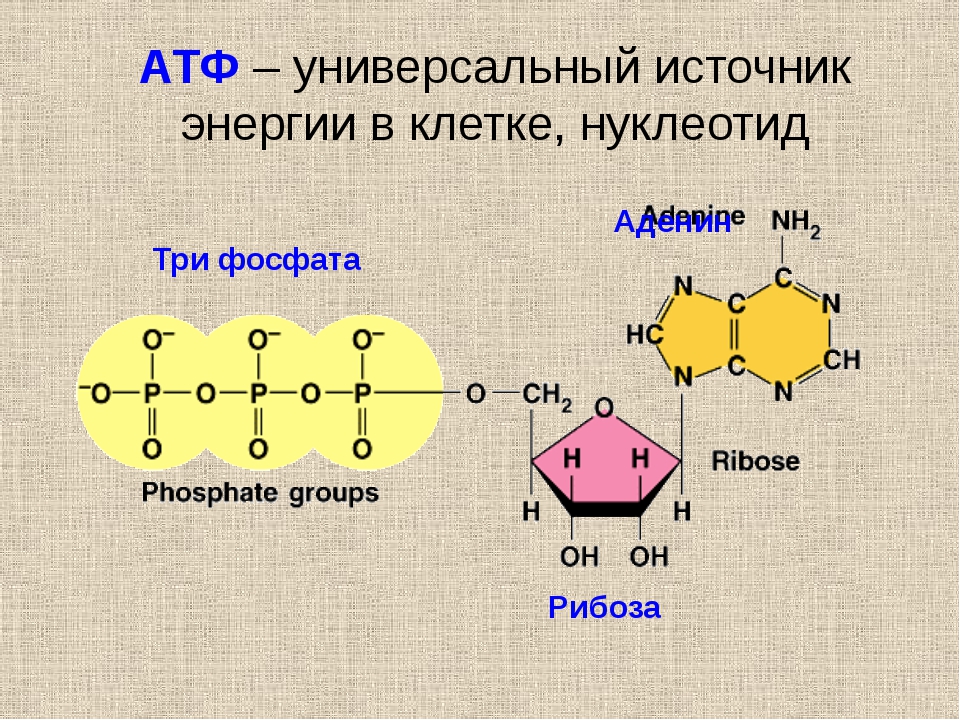
У эукариот митохондриальный FO образует димеры, изгибающие член. Эти димеры самоорганизуются в длинные ряды на концах крист , возможно, это первая стадия образования крист. Атомная модель димерной области дрожжевого F O была определена с помощью крио-ЭМ с общим разрешением 3,6 Å.
Модель привязки
Механизм АТФ-синтазы. Показанные АДФ и P i (розовый) объединены в АТФ (красный), а вращающаяся субъединица γ (гамма), выделенная черным цветом, вызывает конформационные изменения.В 1960–1970-х годах Пол Бойер , профессор Калифорнийского университета в Лос-Анджелесе , разработал теорию механизма изменения связывания или триггера, которая постулировала, что синтез АТФ зависит от конформационного изменения АТФ-синтазы, генерируемого вращением гамма-субъединицы. Исследовательская группа Джона Э. Уокера , работавшего тогда в лаборатории молекулярной биологии MRC в Кембридже , кристаллизовала каталитический домен F 1 АТФ-синтазы.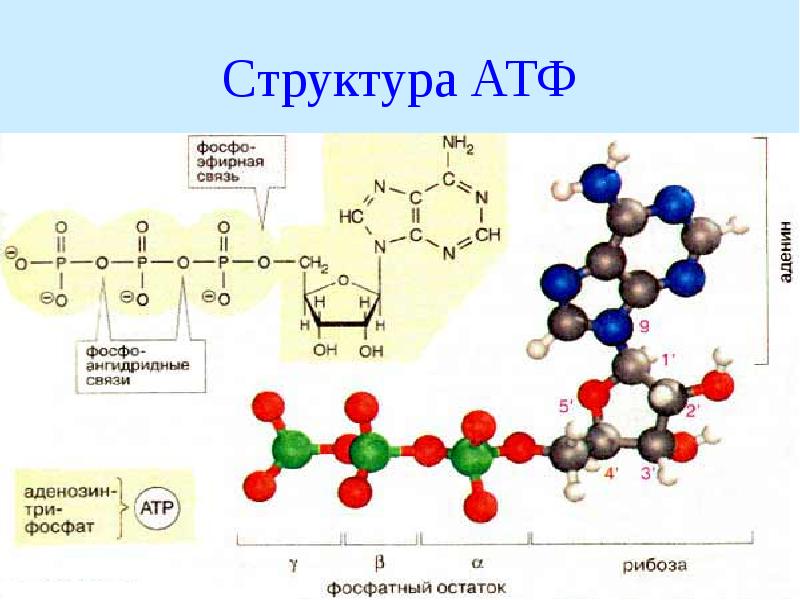 Эта структура, в то время самая большая из известных асимметричных белковых структур, указала на то, что модель роторного катализа Бойера была, по сути, правильной. За разъяснение этого Бойер и Уокер разделили половину Нобелевской премии по химии 1997 года .
Эта структура, в то время самая большая из известных асимметричных белковых структур, указала на то, что модель роторного катализа Бойера была, по сути, правильной. За разъяснение этого Бойер и Уокер разделили половину Нобелевской премии по химии 1997 года .
Кристаллическая структура F 1 показывает чередующиеся альфа- и бета- субъединицы (по 3 каждой), расположенные как сегменты апельсина вокруг вращающейся асимметричной гамма-субъединицы. Согласно современной модели синтеза АТФ (известной как чередующаяся каталитическая модель), трансмембранный потенциал, создаваемый протонными катионами (H +), поставляемыми цепью переноса электронов, перемещает катионы протонов (H +) из межмембранного пространства через мембрану через мембрану. F 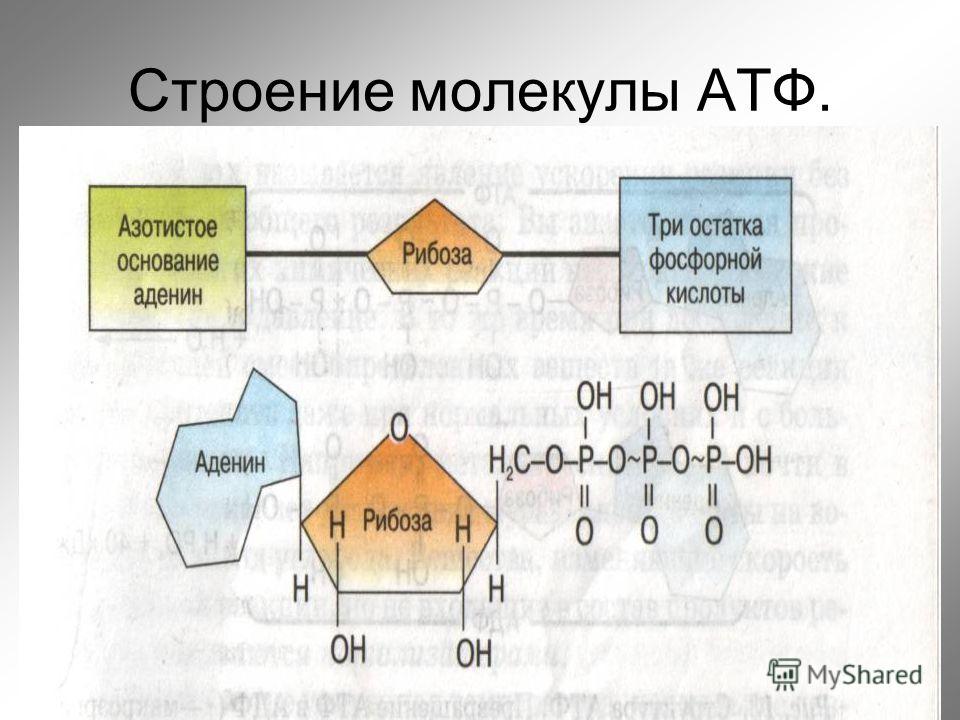 Основные F 1 субъединицы предохранены от вращения в полном согласии с центральной ножкой ротором периферийного стеблом , который присоединяется к альфа — 3 беты 3 к невращающейся части F O . Структура интактной АТФ-синтазы в настоящее время известна с низким разрешением из исследований комплекса с помощью электронной криомикроскопии (крио-ЭМ). Крио-ЭМ модель АТФ — синтазы показывает , что периферийная Стебель представляет собой гибкую структуру , которая оборачивается вокруг комплекса , как он присоединяется к F 1 до F O . При правильных условиях ферментативная реакция также может протекать в обратном направлении, когда гидролиз АТФ запускает перекачку протонов через мембрану.
Основные F 1 субъединицы предохранены от вращения в полном согласии с центральной ножкой ротором периферийного стеблом , который присоединяется к альфа — 3 беты 3 к невращающейся части F O . Структура интактной АТФ-синтазы в настоящее время известна с низким разрешением из исследований комплекса с помощью электронной криомикроскопии (крио-ЭМ). Крио-ЭМ модель АТФ — синтазы показывает , что периферийная Стебель представляет собой гибкую структуру , которая оборачивается вокруг комплекса , как он присоединяется к F 1 до F O . При правильных условиях ферментативная реакция также может протекать в обратном направлении, когда гидролиз АТФ запускает перекачку протонов через мембрану.
Механизм изменения связывания включает в себя циклическое переключение активного сайта субъединицы β между тремя состояниями. В «рыхлом» состоянии АДФ и фосфат попадают в активный центр; на соседней диаграмме это показано розовым цветом. Затем фермент претерпевает изменение формы и объединяет эти молекулы, при этом активный центр в результирующем «плотном» состоянии (показано красным) связывает вновь образованную молекулу АТФ с очень высоким сродством . Наконец, активный сайт возвращается в открытое состояние (оранжевый), высвобождая АТФ и связывая больше АДФ и фосфата, готовый к следующему циклу производства АТФ.
Наконец, активный сайт возвращается в открытое состояние (оранжевый), высвобождая АТФ и связывая больше АДФ и фосфата, готовый к следующему циклу производства АТФ.
Физиологическая роль
Как и другие ферменты, активность АТФ-синтазы F 1 F O обратима. Достаточно большие количества АТФ заставляют его создавать трансмембранный протонный градиент , который используется ферментацией бактерий, у которых нет цепи переноса электронов, а, скорее, гидролизуют АТФ, чтобы создать градиент протонов, который они используют для управления жгутиками и транспорта питательные вещества в клетку.
При дыхании бактерий в физиологических условиях АТФ-синтаза, как правило, работает в противоположном направлении, создавая АТФ, используя движущую силу протона, создаваемую цепью переноса электронов в качестве источника энергии. Общий процесс создания энергии таким образом называется окислительным фосфорилированием . Тот же процесс происходит в митохондриях , где АТФ-синтаза расположена во внутренней митохондриальной мембране, а F 1 -часть проецируется в митохондриальный матрикс .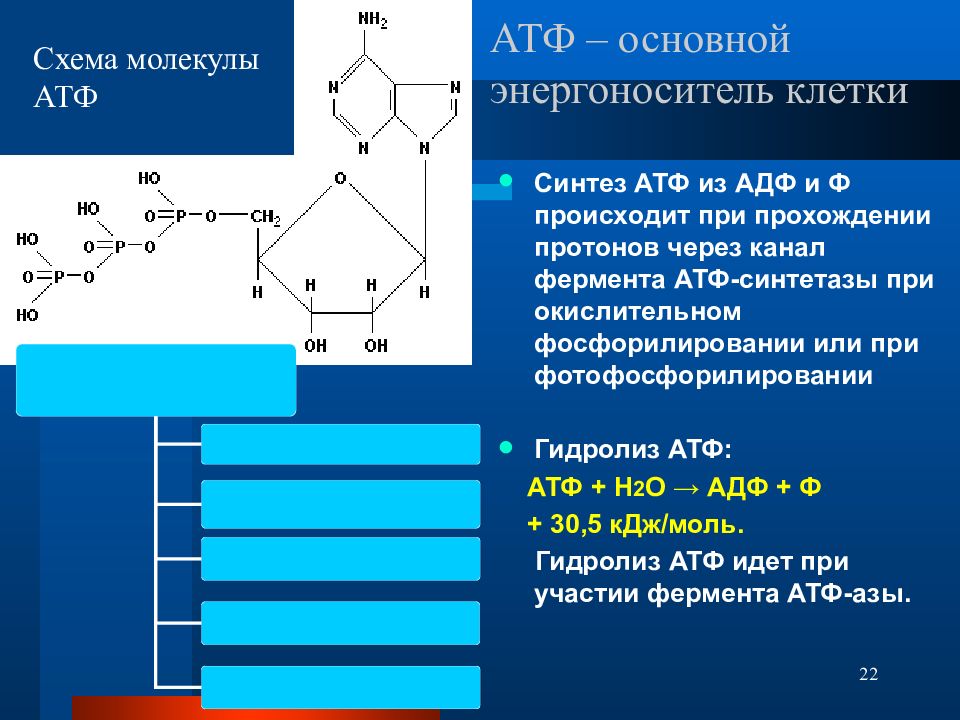 Потребление АТФ АТФ-синтазой закачивает катионы протонов в матрицу.
Потребление АТФ АТФ-синтазой закачивает катионы протонов в матрицу.
Эволюция
Эволюция АТФ — синтазы , как полагают, был модульным , при которой два функционально независимых подразделения стали ассоциироваться и приобрел новые функциональные возможности . Эта ассоциация, по-видимому, возникла на раннем этапе эволюционной истории, потому что по существу одинаковая структура и активность ферментов АТФ-синтазы присутствуют во всех царствах жизни. Синтаза F-АТФ демонстрирует высокое функциональное и механистическое сходство с V-АТФазой . Однако, в то время как F-АТФ-синтаза генерирует АТФ, используя протонный градиент, V-АТФаза генерирует протонный градиент за счет АТФ, генерируя значения pH всего до 1.
Область F 1 также демонстрирует значительное сходство с гексамерными ДНК-геликазами (особенно с фактором Rho ), а вся область фермента показывает некоторое сходство с H+
Приведен T3SS или жгутик моторных комплексов. Гексамер α 3 β 3 области F 1 демонстрирует значительное структурное сходство с гексамерными ДНК-геликазами; оба образуют кольцо с 3-кратной вращательной симметрией с центральной порой. У обоих есть роли, зависящие от относительного вращения макромолекулы в поре; ДНК-геликазы используют спиралевидную форму ДНК, чтобы управлять своим движением вдоль молекулы ДНК и обнаруживать сверхспирали, тогда как α 3 β 3 гексамер использует конформационные изменения через вращение субъединицы γ для запуска ферментативной реакции.
У обоих есть роли, зависящие от относительного вращения макромолекулы в поре; ДНК-геликазы используют спиралевидную форму ДНК, чтобы управлять своим движением вдоль молекулы ДНК и обнаруживать сверхспирали, тогда как α 3 β 3 гексамер использует конформационные изменения через вращение субъединицы γ для запуска ферментативной реакции.
H+
двигатель частицы F O показывает большое функциональное сходство с H+
двигатели, приводящие в движение жгутики. Оба имеют кольцо из множества маленьких альфа-спиральных белков, которые вращаются относительно соседних неподвижных белков, используя H+
градиент потенциала как источник энергии. Однако эта связь незначительна, так как общая структура жгутиковых моторов намного сложнее, чем у частицы F O, а кольцо с примерно 30 вращающимися белками намного больше, чем спиральные белки из 10, 11 или 14 в F O. сложный. Однако более свежие структурные данные показывают, что кольцо и стебель структурно подобны частице F 1 .
Теория модульной эволюции происхождения АТФ-синтазы предполагает, что две субъединицы с независимой функцией, ДНК-геликаза с АТФазной активностью и H+
motor, были способны связываться, и вращение двигателя приводило к обратному развитию АТФазной активности геликазы. Затем этот комплекс приобрел большую эффективность и в конечном итоге превратился в современные сложные АТФ-синтазы. В качестве альтернативы ДНК-геликаза / H+
моторный комплекс мог иметь H+
насосная активность с АТФазной активностью геликазы, управляющей H+
мотор в обратном направлении. Это могло развиться, чтобы осуществить обратную реакцию и действовать как АТФ-синтаза.
Ингибиторы
Было обнаружено множество природных и синтетических ингибиторов АТФ-синтазы. Они использовались для исследования структуры и механизма АТФ-синтазы. Некоторые могут иметь терапевтическое применение. Существует несколько классов ингибиторов АТФ-синтазы, включая пептидные ингибиторы, полифенольные фитохимические вещества, поликетиды, оловоорганические соединения, полиеновые производные α-пирона, катионные ингибиторы, аналоги субстратов, модификаторы аминокислот и другие различные химические вещества. Некоторые из наиболее часто используемых ингибиторов АТФ-синтазы — олигомицин и DCCD .
Существует несколько классов ингибиторов АТФ-синтазы, включая пептидные ингибиторы, полифенольные фитохимические вещества, поликетиды, оловоорганические соединения, полиеновые производные α-пирона, катионные ингибиторы, аналоги субстратов, модификаторы аминокислот и другие различные химические вещества. Некоторые из наиболее часто используемых ингибиторов АТФ-синтазы — олигомицин и DCCD .
У разных организмов
Бактерии
АТФ-синтаза E. coli представляет собой простейшую из известных форм АТФ-синтазы с 8 различными типами субъединиц.
Бактериальные F-АТФазы могут иногда действовать наоборот, превращая их в АТФазу. Некоторые бактерии не имеют F-АТФазы, используя АТФазу A / V-типа двунаправленно.
Дрожжи
АТФ-синтаза дрожжей — одна из наиболее изученных эукариотических АТФ-синтаз; и пять F 1 , восемь субъединиц F O и семь ассоциированных белков были идентифицированы. Большинство этих белков имеют гомологи у других эукариот.
Растение
У растений АТФ-синтаза также присутствует в хлоропластах (CF 1 F O -ATP-синтаза). Фермент интегрирован в тилакоидную мембрану; часть CF 1 прилипает к строме , где происходят темные реакции фотосинтеза (также называемые независимыми от света реакциями или циклом Кальвина ) и синтеза АТФ. Общая структура и каталитический механизм АТФ-синтазы хлоропластов почти такие же, как и у бактериального фермента. Однако в хлоропластах движущая сила протонов создается не дыхательной цепью переноса электронов, а первичными фотосинтетическими белками. Синтаза имеет вставку из 40 аминокислотных остатков в гамма-субъединицу для подавления расточительной активности в темноте.
Млекопитающее
АТФ-синтаза, выделенная из митохондрий сердца крупного рогатого скота ( Bos taurus ), с точки зрения биохимии и структуры является наиболее охарактеризованной АТФ-синтазой. Говяжье сердце используется в качестве источника фермента из-за высокой концентрации митохондрий в сердечной мышце. Их гены имеют близкую гомологию с АТФ-синтазами человека.
Их гены имеют близкую гомологию с АТФ-синтазами человека.
Гены человека, кодирующие компоненты АТФ-синтаз:
- ATP5A1
- ATP5B
- ATP5C1 , ATP5D , ATP5E , ATP5F1 , ATP5G1 , ATP5G2 , ATP5G3 , ATP5H , ATP5I , ATP5J , ATP5J2 , ATP5L , ATP5O
- МТ-АТФ6 , МТ-АТФ8
Другие эукариоты
Эукариоты, принадлежащие к некоторым дивергентным линиям, имеют особую организацию АТФ-синтазы. Эвгленозои АТФ — синтазы образует димер с бумеранг-образной формой F 1 головкой , как и другими митохондриальной АТФ — синтазы, а Р О подкомплексе имеет много уникальных субъединиц. Он использует кардиолипин . Ингибирующий IF- 1 также связывается по-разному, как и трипаносоматида .
Археи
Обычно у архей нет F-АТФазы. Вместо этого они синтезируют АТФ с использованием А-АТФазы / синтазы, вращающейся машины, структурно подобной V-АТФазе, но в основном функционирующей как АТФ-синтаза. Считается, что, как и бактериальная F-АТФаза, она также действует как АТФаза.
Смотрите также
Ссылки
дальнейшее чтение
внешняя ссылка
Онлайн урок: Нуклеиновые кислоты. АТФ. Витамины по предмету Биология 9 класс
Нуклеотидный состав ДНК в 1905 г. впервые количественно проанализировал американский биолог Эрвин Чаргафф.
Он обнаружил, что в молекуле ДНК число пуриновых оснований всегда равно числу пиримидиновых.
Молекулярное количество аденина равно количеству тимина, а количество гуанина равно цитозину- это правило Чаргаффа или принцип комплементарности (дополнительности).
Согласно принципу комплементарности можно восстановить недостающую цепь ДНК.
Задача:
Первая цепочка ДНК имеет следующую последовательность нуклеотидов:
А- Г- Ц- Т- Т- Ц- Г- Г- А- Г
достойте вторую цепочку ДНК, используя принцип комлементарности.
Решение:
Мы видим, что первый нуклеотид в первой цепи ДНК- аденин (А), смотрим правило комплементарности:
значит, аденину (А) соответствует тимин (Т).
Далее второй нуклеотид в первой цепи гуанин (Г). Опять обращаемся к принципу комплементарности, гуанин (Г) соответствует цитозину (Ц).
И таким образом, мы можем достроить всю вторую цепь ДНК.
Первая цепь ДНК: А- Г- Ц- Т- Т- Ц- Г- Г- А- Г
Вторая цепь ДНК: Т- Ц- Г- А- А- Г- Ц- Ц- Т- Ц
Кроме достраивания цепей ДНК в ЕГЭ присутствуют задачи на определение количества (%) нуклеотидов в гене и определение длины гена.
Для решения таких задач тоже используют правило Чаргаффа: молекулярное количество аденина равно количеству тимина, а количество гуанина равно цитозину.
Нуклеотиды расположены на расстоянии друг от друга 0,34 нм и молекулярная масса одного нуклеотида равна 345. Эти величины постоянные, они также используются для решения задач по ДНК.
Примеры задач:
Задача
В молекуле ДНК доля тиминовых нуклеотидов составляет 15% от общего количества нуклеотидов.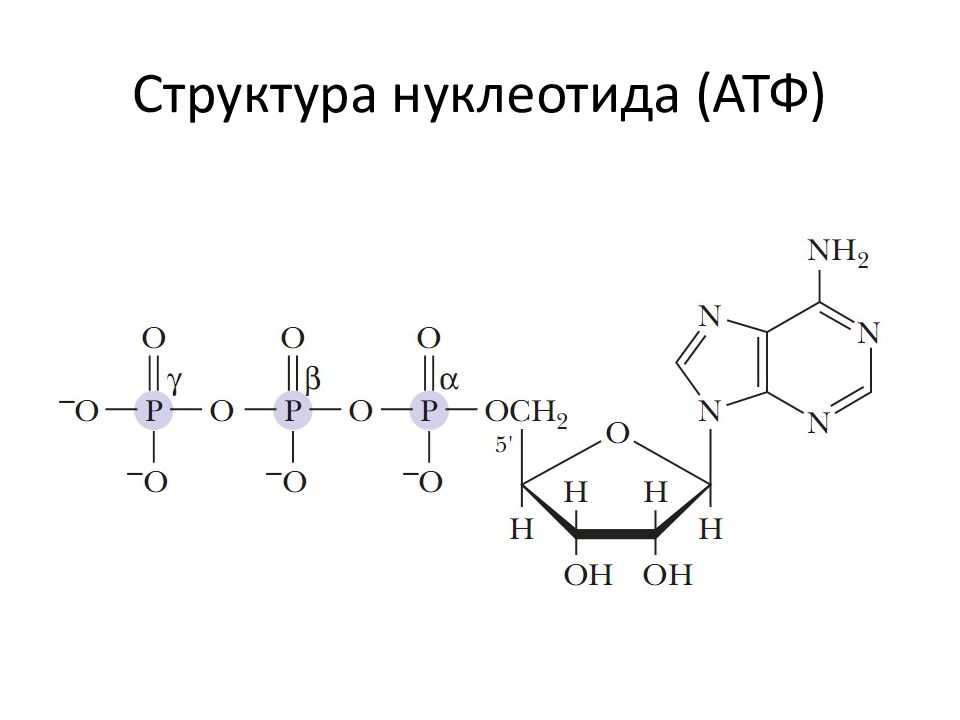
Определите количество других видов нуклеотидов в данной молекуле ДНК.
Решение:
1. По правилу Чаргаффа количество Тимина (Т) в ДНК равно аденину (А), следовательно, если доля Т = 15%, значит, и А будет = 15%.
2. В сумме А + Т = 30%
3. Всего всех нуклеотидов ДНК = 100%, из них на долю А + Т приходится 30%
4. 100% — 30% = 70%, то есть 70% приходится на гуанин (Г) и цитозин (Ц)
5. Количество Ц = Г , следовательно, 70% : 2 = 35% (35% = Г; 35% = Ц)
Ответ: А = (15%), Т = (15%), Г = (35%), Ц = (35%)
Задача
Участок цепи ДНК содержит 1500 нуклеотидов. В одной из цепей содержится 150 нуклеотидов А, 200 нуклеотидов Т, 250 нуклеотидов Г и 150 нуклеотидов Ц. Сколько нуклеотидов каждого вида будет во второй цепи ДНК?
Сколько нуклеотидов каждого вида будет во второй цепи ДНК?
Решение:
По правилу Чаргаффа в ДНК количество гуанина (Г) равно цитозину (Ц), количество тимина (Т) равно аденину (А). Если А в первой цепочке 150 нуклеотидов, значит и Т во второй цепи будет тоже 150, следовательно, получается:
1-я цепь: 2-я цепь:
А = 150 Т = 150
Т = 200 А = 200
Г = 250 Ц = 250
Ц =1 50 Г = 150
Ответ: Во второй цепи ДНК: Т=150; А=200; Ц=250; Г=150
Задача
В молекуле ДНК обнаружено 880 гуаниловых нуклеотидов, которые составляют 22% от общего количества нуклеотидов этой ДНК. Сколько каждого нуклеотида содержится в этой молекуле ДНК? Какова длина этой молекулы ДНК?
Сколько каждого нуклеотида содержится в этой молекуле ДНК? Какова длина этой молекулы ДНК?
Решение:
1) Исходя из принципа комплементарности (А + Т) + (Г+ Ц) = 100%
Тогда количество цитидиловых нуклеотидов равно: Г = Ц = 880, или 22%, то есть Г = 22% и Ц = 22%
2) На долю (Т + А) приходится: 100% — (22% + 22%) = 56%- количество Т и А
3) Необходимо посчитать количество нуклеотидов, исходя из процентных данных. Составляем пропорцию:
880 = 22%
Х = 56%
Х = (880*56) : 22 = 2400 нуклеотидов, приходится в сумме на А+Т
Так как А = Т, то 2400 : 2=1120 нуклеотидов, то есть 1120 = А и 1120 нуклеотидов Т
3) Всего в этой молекуле ДНК содержится (880 х 2) + (1120 х 2) = 4000 нуклеотидов.
4) Для определения длины ДНК узнаем, сколько нуклеотидов содержится в одной цепи:
4000 : 2 = 2000
Мы знаем, что нуклеотиды расположены на расстоянии друг от друга 0,34 нм и вычисляем длину ДНК в одной цепи:
0,34 нм х 2000 нуклеотидов= 680 нм.
Ответ: в молекуле ДНК Г = Ц = 880 и А = Т = 1120 нуклеотидов; длина этой молекулы 680 нм.
Синтез ДНК
Каждая молекула ДНК способна к самоудвоению, в основе которого лежит тот же принцип комлементарности (дополнительности). Этот принцип поможет понять, как строится новая молекула ДНК в новой клетке.
Перед каждым делением клетки (в интерфазе) происходит образование новой молекулы ДНК под действием фермента дезоксирибонуклеазы.
Фермент разрывает двойную цепь ДНК и спираль раскручивается.
Каждая отдельная цепь собирает новую молекулу ДНК по принципу комплементарности, в результате образуется две молекулы ДНК.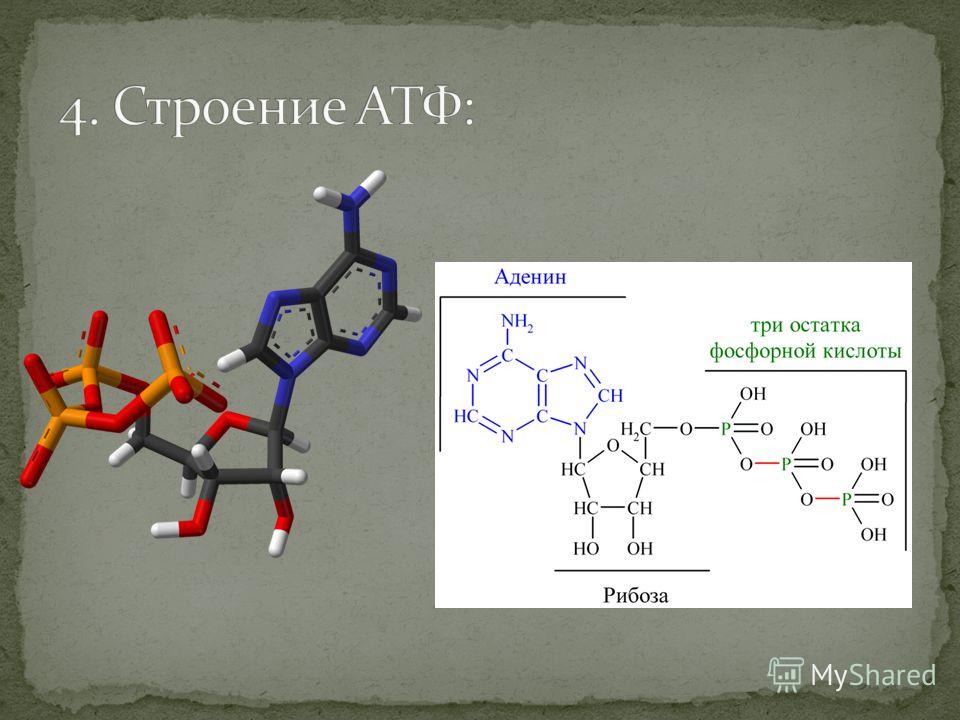
Этот процесс называется редупликация ДНК— копирование молекулы ДНК.
Руководство по структуре ATF
Стив Тинни
Версия от 13.10.2019
Этот документ дает руководство по вводу особенности блочной структуры текстов в ATF, транслитерация ASCII Формат.
Мы предлагаем простое введение в набор ATF тексты, сначала описывая наиболее общие функции и заполняя подробности позже. Прежде чем объяснять какие-либо особенности, вот простой типичный пример текста ATF:
& P555555 = Некоторая публикация 32 @лицевой 1.1 (диш) уду ба-угу $ обратный пустой
В этом примере показаны четыре наиболее распространенных типа линий. в тексте ATF:
- и линии
- Каждый текст ATF должен начинаться с
& -lines(«and-lines»), который обычно дает P-идентификатор CDLI и должен также иметь удобочитаемое имя после знака ‘=’.
- @ -строчный
- Разделы в тексте указываются с помощью строк, начинающихся с
Знак
@(«at-lines»).Они используются для обозначения объекта типы, поверхности, подразделения и колонны. - $ — линии
- Пояснительные замечания относительно сохранности или состояния
текст дается с использованием
$ -строк(«долларовых строк»). Эти выглядят как обычные последовательности слов, но могут быть подвержены строгие правила. - Текстовые строки
- Строки, начинающиеся с пробелов, за которыми следует точка, за которой следует один или несколько пробелов — это строки текста. Правила транслитерации даны в графеме ATF руководство.
Другой довольно распространенный тип строки в файле ATF начинается с
знак решетки ( # ). Есть два типа # -line: протоколы
и комментарии.
Протоколы
Протоколы — это утверждения, которые интерпретируются или сохраняются ATF. процессора, но не являются частью собственно текстового издания. Протоколы все названы и могут запускать специальную обработку в ATF процессор.
Протоколы указываются в ATF строкой, начинающейся с хеша
символ ( # ), известное имя протокола и символ двоеточия
(: ).
Подробная информация о протоколах выходит за рамки этого руководства; за теперь достаточно знать, что они выглядят так:
& P123321 = Немного аккадского текста
#atf: lang akk-x-oldbab
1. i-na AN {+ e}
#note: Это надуманная заметка. Большинство протоколов представляют собой одну строку и не требуют
пустая строка после них, чтобы отделить их от следующего протокола
(единственное исключение — #note: ).
Дополнительная информация о протоколах, что это такое, где они находятся разрешено, а правила заказа протоколов доступны в руководство по протоколам.
Комментарии
Комментарии — это отступления, которые не являются частью текстового издания или аннотация; они полезны для хранения нечетных битов информации в файл, не мешая текстовой редакции или аннотация.
Комментарии указываются в ATF одной или несколькими строками, начинающимися с
символ решетки ( # ).
Комментарии выглядят как протоколы в том смысле, что начинаются с решетки, но они не могут начинаться с последовательности хэш-имя-двоеточие.Комментарии могут быть включены в транслитерацию текста, но не раньше первого текст в файле. Комментарии всегда должны следовать любым протоколам, которые происходят рядом с ними.
Последовательность строк, начинающаяся с решетки, представляет собой многострочную комментарий. Чтобы разделить несколько комментариев в одной строке, используйте пробел строка в файле ATF.
1. а # простой комментарий 2. а # более длинный комментарий, который несколько искусственно расширяется # над несколькими строками 3. а # один комментарий к строке 3. # еще один комментарий к строке 3.4. а # Комментарии немного похожи на протоколы, но нет никаких #confusion: об этом позаботятся правила сканирования процессора ATF. ).Вы также можете указать, что примечание соответствует метке текстовой строки (или диапазона) с помощью обозначение@notelabel {...}, например,#note: @notelabel {i 1} Примечание к строке первого столбца 1..& -строчки используются для введения нового текста и состоят из двух части: идентификатор и имя.
Для транслитерации экземпляров идентификатор - это буква "P", за которой следует шесть цифры, например, P123456. Этот идентификатор назначается CDLI и является ссылочный ID объекта в основном каталоге CDLI; получить идентификаторы для объекты, не входящие в каталог CDLI, отправьте электронное письмо по адресу cdli @ cdli.ucla.edu.
Название текста должно совпадать с «Обозначением». поле в основном каталоге CDLI; процессор ATF обнаруживает несовпадения и сообщает правильное имя. Этот механизм предназначен для обеспечения убедитесь, что P-номер в идентификаторе действительно ссылается на текст транслитератор намеревается.
В ATF две части & -линии разделены пространство-равно-пространство, например:
& P000001 = ATU 3, пл. 011, W 6435, а@ -line используются для структурных тегов.Несколько с помощью этого механизма можно указать виды структур: физические структура, например, объекты, поверхности; структура рукописи, т.е. колонны; и структура документа, например, разделы и колофоны. За ясности, мы описываем здесь только структурные особенности, которые разрешены в транслитерации объекта, т.е. тексты с идентификатором, начинающимся с
P. Документация структурных соглашений для составные тексты даны в композитах руководство.Объектов
Вид объекта, на котором транслитерируется надпись. написанное обозначается одним из следующих тегов:
-
@ планшет - По умолчанию и, следовательно, необязательно; объект - планшет.
-
@ конверт - Таблетки и конверты с одинаковым номером P можно транслитерировать отдельно используя этот тег.
-
@ призма - Объект - призма.
-
@bulla - Объект - булла.
-
@ фрагмент - Объект - это фрагмент с именем фрагмента (например, буква)
после тега; может использоваться более одного раза для транслитерации
несколько фрагментов объекта, например:
& P212121 = Какой-то фрагментарный объект @ фрагмент а 1.а @ фрагмент b 1. а
-
@ объект - Общий тег объекта, за которым должен следовать тип
объект, например
@object Каменный парик.
Уплотнения
Транслитерация текста, начертанного на объекте физической печати.
следует обрабатывать с помощью тега @object :
& P333444 = Некоторая печать @объект печать 1. да-да 2. dumu du-du
Поверхности
Поверхности - это в основном физические поверхности:
-
@обверс,@ обратный - Аверс и реверс.
-
@ слева,@ справа,@ вершина,@ снизу - Задаваемые края, левый, правый, верхний и нижний (как видно при глядя на лицевую сторону планшета).
-
@ лицо - Условное обозначение поверхностей призмы; должен соблюдаться
одной строчной буквой, обозначающей лицо, например:
& P123321 = Немного призмы @ призма @face a 1. а @face b 1. e
-
@ поверхность - Общий тег поверхности, за которым должно следовать имя поверхности,
е.г .:
@ поверхностное плечо;@ поверхностная сторона. -
@ край - Базовая метка края; может сопровождаться одной строчной буквой
назвать край аналогично
@face.
Уплотнения
Транслитерация пломбы должна выполняться с использованием @seal тег включен как поверхность после
транслитерация объекта, на котором происходит пломбирование:
& P343434 = Запечатанная табличка 1.а $ печать 1 @ печать 1 1. du-du
Использование печати $ предвосхищает обсуждение
$ -строчки ниже; этот механизм можно использовать, чтобы указать, какие уплотнения
произойти где на объекте.
Колонны
Столбцы обозначаются тегом @column , который может
опускаться для текстов в одну колонку. Номера столбцов должны быть указаны в
арабские цифры:
& P545454 = Некоторый текст в столбцы @column 1 1. а @column 2 1. e
Статус
Состояние некоторых функций, обозначенных @-линиями, может быть обозначены аналогично графемам; обозначение должно быть естественным и следовать ассириологическим соглашениям:
@лицевой?
Значение: состояние аверса / реверса неопределенное
@reverse! *
Значение: с подборкой; обратное правильно, несмотря на обозначение в публикации
Простые числа могут использоваться там, где это имеет смысл:
@face a ' @column 3 '
Заголовки
Транслитерация и составные части могут содержать заголовки, которые имеют вид @h , где DIGIT - это уровень структуры заголовка, обычно 1, 2 или 3.
Вехи
По техническим причинам невозможно переплетение физических структура (описанная выше для транслитерированных объектов) и структура документа (например, разделение на абзацы). Это ограничение решается путем обращения к вехам.
Подразделения
Документарные подразделения в транслитерированном объекте задаются с использованием
тег @m с типом вехи, указанным после знака равенства
знак и следующий вид деления; необязательное название подразделения или
номер может следовать за типом деления:
@ m = разделение абзаца 1 @ m = деление колофона
Дискурс
Простая поддержка элементов дискурса в административных и научных тексты предоставляются с использованием сокращений, которые также реализованы как вехи.Эти сокращения:
-
@catchline -
@colophon -
@ дата -
@ подписии@ подпись -
@сводка -
@ очевидцы
Эти вехи должны быть указаны между строками. Если вам нужно отметить веху в середине строки, тогда вы можете разделить строку на две (помечены, например, a и b) на этапе.
& P787878 = Некоторый административный текст 1.1 (диш) уду 2. да-да 3. šu ba-ti @Дата 4. u₄ 1-kam @осталось @резюме 1. 1 (диш) уду
& P8 = Научный текст
@colophon
1а. UNUG {ki}
@Дата
1b. {iti} AB U₄ 1-KAM₂
2. MU 1.39@v-KAM₂ {m} an-ti-ʾi-ku-su LUGAL
$ -строчки используются для обозначения информации о состоянии текст или объект, или описать особенности объекта, которые не часть собственно транслитерации. Они бывают двух видов: строгие и свободно.
Strict $ -строки подчиняются ограничениям в таблица ниже; строгие $ -строки можно интерпретировать полностью процессором ATF, и интерпретированная информация может затем использоваться другими программами.Лучше всего использовать строгие $-строки.
Свободные $ - строки обозначаются скобками вокруг содержимого $ -line. Это средство, предоставленное включить аннотацию функций, на которые не распространяется строгий $ -строчная спецификация. Если процессор ATF обнаруживает, что $ -line фактически соответствует критериям, определенным для строгих $-линий, которые он дает информационное уведомление о том, что скобки следует удалить.
$ -строки и комментарии - это два совершенно разных средства, но опыт показал, что транслитераторы могут спутать эти два понятия.Комментарии предназначены для информации, не относящейся к транслитерация и описание текста; комментарии не отображается, когда текст отформатирован для отображения или печати. $ -линии предназначены для информации, которая является неотъемлемой частью понимания текстовые данные; $ -lines включаются, когда текст отображается или напечатан.
Уплотнение
В частности, символы $ -lines используются для обозначения того, что печать используется на объект; форма:
$ печать
Где N - число, указывающее, какая печать используется;
если транслитерация печати также дается с использованием
Заголовок @seal , номер после $ seal должен соответствовать номеру после @seal .Увидеть
пример выше.
Состояние
Большинство $-строк используются для предоставления информации о состоянии транслитерируемый объект. Условные обозначения для этого могут быть резюмируется следующим образом:
| Квалификация | Объем 1 | Область | Состояние | |
|---|---|---|---|---|
1 Степень N может быть
число, такое как 1 или 5; ДИАПАЗОН дает два числа
разделенные дефисом, e.г., 3-5. | ||||
2 ОБЪЕКТ - любой объект
спецификатор, как описано выше, например, планшет, объект и т. д. | ||||
3 ПОВЕРХНОСТЬ - любая поверхность
спецификатор, как описано выше, например, аверс, левый и т. д. | ||||
| не менее не более около | n несколько некоторые НОМЕР ДИАПАЗОН остаток начало начало середина конец | ОБЪЕКТ 2 ПОВЕРХНОСТЬ 3 столбец столбца строка строки корпус корпуса поверхность | пусто сломано стерто неразборчиво отсутствует следов | |
Постановления
Строки$ также используются для обозначения заслуживающих внимания постановлений по планшет; обычное решение по делу или строке не должно обозначаться $ -line, но там, где писец использовал правило, чтобы дать дополнительные информацию о структуре документа следует указать как:
(одинарная | двойная | тройная) постановка
Примеры
Строгие $ -строки выглядят так:
$ 3 пустые строки $ Остальная часть аверса отсутствует
Свободная линия $ выглядит так:
$ (голова статуи сломана)
Управляющая строка $ выглядит так:
$ двойное решение
Изображения
Встроенные изображения можно указать с помощью формы:
$ (изображение N = <текст>)
Где N - номер изображения, состоящий из цифр, за которыми следует
необязательные строчные буквы от a до z, а ).
$ (изображение 1 = нумерованная диаграмма треугольника)
В настоящее время реализация работает только для XHTML, который
произведено в рамках проекта. Процессор ATF создает имя файла
состоящий из идентификатора текста и значения N изображения, соединенных
знак (например, P123456 @ 1 ). Затем производитель XHTML излучает тег с атрибутом
src , установленным на /.
Таким образом, в настоящей реализации должен существовать
файл с соответствующим именем в графическом формате PNG, находящийся в
каталог изображений проекта .Реализация
ожидается, что он будет поддерживать более сложный механизм локатора в
будущее.
Строки транслитерированного текста начинаются с последовательности непробелов. символы, за которыми следует точка и пробел (обычно это номера, но это не обязательно):
1. а а + 1. е 2 '. я
В ATF игнорируются строки, содержащие только пробелы; линии начинающиеся с пробела - это строки продолжения, новая строка и ведущие пробелы удаляются процессором ATF:
1.а а а а а
Содержание строк определяется в основном языком описания графем, но есть некоторые линейные функции ATF, которые не нужны многим пользователям и которые рассматриваются в расширенных документация.
Вопросы по этому документу можно направлять по адресу Руководящий комитет Oracc (osc at oracc dot org).
Oracc: Открытый богато аннотированный корпус клинописи
Этот документ дает руководство по вводу особенности блочной структуры текстов в ATF, транслитерация ASCII Формат.
Введение
Мы предлагаем простое введение в набор ATF тексты, сначала описывая наиболее общие функции и заполняя подробности позже. Прежде чем объяснять какие-либо особенности, вот простой типичный пример текста ATF:
& P555555 = Некоторая публикация 32 @лицевой 1. 1 (диш) уду ба-угу $ обратный пустой
В этом примере показаны четыре наиболее распространенных типа линий. в тексте ATF:
- и линии
- Каждый текст ATF должен начинаться с
& -lines("and-lines"), который обычно дает P-идентификатор CDLI и должен также иметь удобочитаемое имя после знака '='. - @ -строчный
- Разделы в тексте указываются с помощью строк, начинающихся с
Знак
@("at-lines"). Они используются для обозначения объекта типы, поверхности, подразделения и колонны. - $ - линии
- Пояснительные замечания относительно сохранности или состояния
текст дается с использованием
$ -строк("долларовых строк"). Эти выглядят как обычные последовательности слов, но могут быть подвержены строгие правила. - Текстовые строки
- Строки, начинающиеся с пробелов, за которыми следует точка, за которой следует один или несколько пробелов - это строки текста.Правила транслитерации даны в графеме ATF руководство.
Типы линий
# -lines
Другой довольно распространенный тип строки в файле ATF начинается с
знак решетки ( # ). Есть два типа # -line: протоколы
и комментарии.
Протоколы
Протоколы - это утверждения, которые интерпретируются или сохраняются ATF. процессора, но не являются частью собственно текстового издания. Протоколы все названы и могут запускать специальную обработку в ATF процессор.
Подробная информация о протоколах выходит за рамки этого руководства; за теперь достаточно знать, что они выглядят так:
& P123321 = Немного аккадского текста
#atf: lang akk-x-oldbab
1. i-na AN {+ e}
#note: Это надуманная заметка. Большинство протоколов представляют собой одну строку и не требуют
пустая строка после них, чтобы отделить их от следующего протокола
(единственное исключение - #note: ).
Дополнительная информация о протоколах, что это такое, где они находятся разрешено, а правила заказа протоколов доступны в руководство по протоколам.
Комментарии
Комментарии - это отступления, которые не являются частью текстового издания или аннотация; они полезны для хранения нечетных битов информации в файл, не мешая текстовой редакции или аннотация.
Комментарии указываются в ATF одной или несколькими строками, начинающимися с
символ решетки ( # ).
Комментарии выглядят как протоколы в том смысле, что начинаются с решетки, но они не могут начинаться с последовательности хэш-имя-двоеточие.Комментарии могут быть включены в транслитерацию текста, но не раньше первого текст в файле. Комментарии всегда должны следовать любым протоколам, которые происходят рядом с ними.
Последовательность строк, начинающаяся с решетки, представляет собой многострочную комментарий. Чтобы разделить несколько комментариев в одной строке, используйте пробел строка в файле ATF.
1. а # простой комментарий 2. а # более длинный комментарий, который несколько искусственно расширяется # над несколькими строками 3. а # один комментарий к строке 3. # еще один комментарий к строке 3.4. а # Комментарии немного похожи на протоколы, но нет никаких #confusion: об этом позаботятся правила сканирования процессора ATF. ).Вы также можете указать, что примечание соответствует метке текстовой строки (или диапазона) с помощью обозначение@notelabel {...}, например,#note: @notelabel {i 1} Примечание к строке первого столбца 1..и линии
& -строчки используются для введения нового текста и состоят из двух части: идентификатор и имя.
Для транслитерации экземпляров идентификатор - это буква "P", за которой следует шесть цифры, например, P123456. Этот идентификатор назначается CDLI и является ссылочный ID объекта в основном каталоге CDLI; получить идентификаторы для объекты, не входящие в каталог CDLI, отправьте электронное письмо по адресу cdli @ cdli.ucla.edu.
Название текста должно совпадать с «Обозначением». поле в основном каталоге CDLI; процессор ATF обнаруживает несовпадения и сообщает правильное имя. Этот механизм предназначен для обеспечения убедитесь, что P-номер в идентификаторе действительно ссылается на текст транслитератор намеревается.
В ATF две части & -линии разделены пространство-равно-пространство, например:
& P000001 = ATU 3, пл. 011, W 6435, а@ -строчный
@ -line используются для структурных тегов.Несколько с помощью этого механизма можно указать виды структур: физические структура, например, объекты, поверхности; структура рукописи, т.е. колонны; и структура документа, например, разделы и колофоны. За ясности, мы описываем здесь только структурные особенности, которые разрешены в транслитерации объекта, т.е. тексты с идентификатором, начинающимся с
P. Документация структурных соглашений для составные тексты даны в композитах руководство.Объектов
Вид объекта, на котором транслитерируется надпись. написанное обозначается одним из следующих тегов:
-
@ планшет - По умолчанию и, следовательно, необязательно; объект - планшет.
-
@ конверт - Таблетки и конверты с одинаковым номером P можно транслитерировать отдельно используя этот тег.
-
@ призма - Объект - призма.
-
@bulla - Объект - булла.
-
@ фрагмент - Объект - это фрагмент с именем фрагмента (например, буква)
после тега; может использоваться более одного раза для транслитерации
несколько фрагментов объекта, например:
& P212121 = Какой-то фрагментарный объект @ фрагмент а 1.а @ фрагмент b 1. а
-
@ объект - Общий тег объекта, за которым должен следовать тип
объект, например
@object Каменный парик.
Уплотнения
Транслитерация текста, начертанного на объекте физической печати.
следует обрабатывать с помощью тега @object :
& P333444 = Некоторая печать @объект печать 1. да-да 2. dumu du-du
Поверхности
Поверхности - это в основном физические поверхности:
-
@обверс,@ обратный - Аверс и реверс.
-
@ слева,@ справа,@ вершина,@ снизу - Задаваемые края, левый, правый, верхний и нижний (как видно при глядя на лицевую сторону планшета).
-
@ лицо - Условное обозначение поверхностей призмы; должен соблюдаться
одной строчной буквой, обозначающей лицо, например:
& P123321 = Немного призмы @ призма @face a 1. а @face b 1. e
-
@ поверхность - Общий тег поверхности, за которым должно следовать имя поверхности,
е.г .:
@ поверхностное плечо;@ поверхностная сторона. -
@ край - Базовая метка края; может сопровождаться одной строчной буквой
назвать край аналогично
@face.
Уплотнения
Транслитерация пломбы должна выполняться с использованием @seal тег включен как поверхность после
транслитерация объекта, на котором происходит пломбирование:
& P343434 = Запечатанная табличка 1.а $ печать 1 @ печать 1 1. du-du
Использование печати $ предвосхищает обсуждение
$ -строчки ниже; этот механизм можно использовать, чтобы указать, какие уплотнения
произойти где на объекте.
Колонны
Столбцы обозначаются тегом @column , который может
опускаться для текстов в одну колонку. Номера столбцов должны быть указаны в
арабские цифры:
& P545454 = Некоторый текст в столбцы @column 1 1. а @column 2 1. e
Статус
Состояние некоторых функций, обозначенных @-линиями, может быть обозначены аналогично графемам; обозначение должно быть естественным и следовать ассириологическим соглашениям:
@лицевой?
Значение: состояние аверса / реверса неопределенное
@reverse! *
Значение: с подборкой; обратное правильно, несмотря на обозначение в публикации
Простые числа могут использоваться там, где это имеет смысл:
@face a ' @column 3 '
Заголовки
Транслитерация и составные части могут содержать заголовки, которые имеют вид @h , где DIGIT - это уровень структуры заголовка, обычно 1, 2 или 3.
Вехи
По техническим причинам невозможно переплетение физических структура (описанная выше для транслитерированных объектов) и структура документа (например, разделение на абзацы). Это ограничение решается путем обращения к вехам.
Подразделения
Документарные подразделения в транслитерированном объекте задаются с использованием
тег @m с типом вехи, указанным после знака равенства
знак и следующий вид деления; необязательное название подразделения или
номер может следовать за типом деления:
@ m = разделение абзаца 1 @ m = деление колофона
Дискурс
Простая поддержка элементов дискурса в административных и научных тексты предоставляются с использованием сокращений, которые также реализованы как вехи.Эти сокращения:
-
@catchline -
@colophon -
@ дата -
@ подписии@ подпись -
@сводка -
@ очевидцы
Эти вехи должны быть указаны между строками. Если вам нужно отметить веху в середине строки, тогда вы можете разделить строку на две (помечены, например, a и b) на этапе.
& P787878 = Некоторый административный текст 1.1 (диш) уду 2. да-да 3. šu ba-ti @Дата 4. u₄ 1-kam @осталось @резюме 1. 1 (диш) уду
& P8 = Научный текст
@colophon
1а. UNUG {ki}
@Дата
1b. {iti} AB U₄ 1-KAM₂
2. MU 1.39@v-KAM₂ {m} an-ti-ʾi-ku-su LUGAL
$ - линии
$ -строчки используются для обозначения информации о состоянии текст или объект, или описать особенности объекта, которые не часть собственно транслитерации. Они бывают двух видов: строгие и свободно.
Strict $ -строки подчиняются ограничениям в таблица ниже; строгие $ -строки можно интерпретировать полностью процессором ATF, и интерпретированная информация может затем использоваться другими программами.Лучше всего использовать строгие $-строки.
Свободные $ - строки обозначаются скобками вокруг содержимого $ -line. Это средство, предоставленное включить аннотацию функций, на которые не распространяется строгий $ -строчная спецификация. Если процессор ATF обнаруживает, что $ -line фактически соответствует критериям, определенным для строгих $-линий, которые он дает информационное уведомление о том, что скобки следует удалить.
$ -строки и комментарии - это два совершенно разных средства, но опыт показал, что транслитераторы могут спутать эти два понятия.Комментарии предназначены для информации, не относящейся к транслитерация и описание текста; комментарии не отображается, когда текст отформатирован для отображения или печати. $ -линии предназначены для информации, которая является неотъемлемой частью понимания текстовые данные; $ -lines включаются, когда текст отображается или напечатан.
Уплотнение
В частности, символы $ -lines используются для обозначения того, что печать используется на объект; форма:
$ печать
Где N - число, указывающее, какая печать используется;
если транслитерация печати также дается с использованием
Заголовок @seal , номер после $ seal должен соответствовать номеру после @seal .Увидеть
пример выше.
Состояние
Большинство $-строк используются для предоставления информации о состоянии транслитерируемый объект. Условные обозначения для этого могут быть резюмируется следующим образом:
| Квалификация | Объем |
|---|
Фотосинтез 1Фотосинтез и клеточное дыханиеДополнения к биологии 101 Cell Unit1.Флуоресценция в растворе хлорофилла
2. Упрощенная иллюстрация митохондрии
3. Структура и функции ATP Структура аденозинмонофосфата, нуклеотида РНК, содержащего пуриновое основание аденин, очень похожа на АТФ (аденозинтрифосфат), за исключением того, что АТФ имеет три фосфата (PO 4 ) вместо одного.АТФ синтезируется во всех живых клетках путем добавления фосфата к АДФ (аденозиндифосфату). АТФ - это молекула жизненно важной энергии всех живых систем, которая абсолютно необходима для ключевых биохимических реакций внутри клеток. Конечный (3-й) фосфат АТФ переносится на другие молекулы клетки, тем самым делая их более реактивными. Например, моносахарид глюкоза очень стабильна при обычных температурах тела и потребует большого количества тепла (например, от пламени), чтобы расщепить ее на диоксид углерода и воду.После получения фосфата из АТФ (процесс, называемый фосфорилированием), глюкоза становится глюкозо-фосфатной и может ферментативно расщепляться в течение нескольких секунд. Большая часть АТФ в эукариотических клетках животных вырабатывается внутри клеточных органелл, называемых митохондриями, в результате окисления глюкозы, процесса, называемого клеточным дыханием. Глюкоза соединяется с кислородом (окисление), образуя углекислый газ, воду и 38 молекул АТФ. В процессе окисления электроны глюкозы перемещаются через железосодержащую ферментную систему цитохрома на внутренних митохондриальных мембранах (называемых кристами).Фактический синтез АТФ путем связывания АДФ (аденозиндифосфата) с фосфатом очень сложен и включает механизм, называемый хемиосмосом. Электронный поток генерирует более высокую концентрацию (заряд) положительно заряженных ионов водорода (H +) (или протонов) на одной стороне мембраны. Когда одна сторона мембраны достаточно «заряжена», эти протоны повторно пересекают мембрану через специальные каналы (поры), содержащие фермент АТФ-синтетазу, поскольку образуются молекулы АТФ.Подробный пошаговый анализ глюкозы во время клеточного дыхания называется циклом Кребса или циклом лимонной кислоты. 4. Упрощенное изображение хлоропласта
Световые реакции фотосинтеза
|
PPT - Структура АТФ Презентация в PowerPoint, скачать бесплатно
Структура АТФ • Каждая молекула АТФ состоит из трех субъединиц: • -Рибозный сахар • -аденин • -Трифосфатная группа, образованная связью трех фосфатные группы (PO4). Как АТФ накапливает, передает и высвобождает энергию • АТФ накапливает энергию, высвобождаемую в результате других экзергонических реакций, в ковалентных связях, которые связывают фосфатные группы в АТФ, называемых фосфоангидридными связями.• Фосфоангидридные связи довольно нестабильны, и для их разрыва требуется лишь небольшое количество свободной энергии. Этот процесс называется гидролизом. • Гидролиз концевой фосфоангидридной связи высвобождает запасенную энергию.
АТФ - это энергетическая валюта клеток • АТФ «связывает» ферментативные реакции высвобождения энергии (экссергонические) и накопления энергии (эндергонические) в клетках. • АТФ, АДФ и фосфат постоянно рециркулируются в живых клетках, поскольку клетки используют групповой перенос для захвата, переноса и высвобождения энергии.
Групповой перенос путем фосфорилирования • Энергия, запасенная в АТФ, легко передается вместе с группой PO4 другим молекулам в процессе, называемом фосфорилированием. • АТФ имеет высокий потенциал группового переноса. АТФ теряет запасенную энергию из-за гидролиза с образованием АДФ и фосфатной группы. Соединение A получает больше энергии за счет фосфорилирования.
Эволюция цыпленка и яйца Какие были первые белки или ДНК? Без ДНК невозможно направить синтез белка, но ДНК не может синтезировать сама себя.Считается, что в первом живом организме РНК (или даже предшественник РНК) была как генетической информацией, так и катализаторами (самореплицирующейся РНК). Сколько реакций катализируют эти РНК-организмы? Считается, что у них была разнообразная биохимия. Считается, что они использовали все основания, содержащие кофакторы и молекулы: НАД, НАДФ, КоА и т. Д. Основание было «ручкой» для этих рибозимов. Если организмы были такими сложными, у них почти наверняка была мембрана. Нет смысла производить НАДН или НАД, если он просто собирается плавать.
Рибозимы Относительно новые открытия • Рибозимы - сегменты РНК, которые проявляют ферментативную активность в отсутствие белка • Примеры: РНКаза P и пептидилтрансфераза • Абзимы - антитела, возникающие для связывания переходного состояния интересующей реакции
Примеры • Рибонуклеаза P (РНКаза P) представляет собой тип рибонуклеазы, которая расщепляет РНК. РНКаза P - это рибозим - рибонуклеиновая кислота, которая действует как катализатор так же, как и фермент на основе белка.Его функция заключается в отщеплении дополнительной или предшественницы последовательности РНК на молекулах тРНК. • Активность пептидилтрансферазы опосредуется не какими-либо рибосомными белками, а рибосомной РНК (рРНК), рибозимом. Этот реликт РНК - наиболее значимое свидетельство, подтверждающее гипотезу мира РНК.
История • 1982: Самосплайсинг в пре-рРНК Tetrahymena (интрон группы I) • Kruger et al, and Cech, Cell 31, 147-157 (1982) • 1983: РНКаза P является рибозимом • Guerrier -Takada et al, и Altman, Cell, 35, 849-857 (1983)
Сколько рибозима? - рибозим в форме головки молотка (вирус растения) - рибозим шпильки (вирус растений) - рибозим дельта гепатита (вирус человека) - рибозим нейроспоры VS (митохондриальная РНК) - рибозим интрона группы I и группы II (рРНК и мт РНК) - РНКаза P (созревание тРНК) - Рибосома (перевод) - Сплайсосома ?? (сплайсинг)
Рибозимы Самосплайсинговые интроны in vivo не являются катализатором, но из них можно сконструировать настоящий катализатор, который функционирует in vitro.Рибозимы очень похожи на белки, и у них есть трехмерная структура, необходимая для их функционирования. Рибозимы измеряют многие из тех же свойств, что и ферменты (kcat, KM и Vmax). Фермент, который катализирует аналогичную реакцию с рибозимом, почти всегда является лучшим катализатором (kcat / KM). У белков больше функциональных групп. Рибозимы медленнее катализируют химию и имеют больше проблем с субстратом / продуктом, связыванием / высвобождением. Существуют также «избранные» ДНК-катализаторы in vitro. Они, как правило, являются более плохими катализаторами, чем рибозимы (2 ’OH играют важную роль в укладке и каталитических свойствах рибозимов).
Каталитическая эффективность, условие - рибозим следует кинетике Михаэлиса-Ментен k1 k2 E + S ES E + P k-1 k-1 + k2 Km = kcat = 0,5-2 мин-1 = 10-5- 10-7 M k1 kcat / Km = 103-106 M-1.min-1 Хорошая каталитическая эффективность !! - все рибозимы нуждаются в катионах для активности (Mg2 +, Mn2 +)
ABZYMES (антитело + ферменты) • Abzymes - антитела, вызванные для связывания переходного состояния интересующей реакции
ABZYMES () Antibody Моноклональные антитела Каталитическая активность Искусственная конструкция Нормальные люди Аутоиммунные заболевания ПРИНЦИП Снижение энергии активации Стабилизация менее благоприятных промежуточных соединений
идея • Используя высокоспецифичные антигенсвязывающие свойства антител, были разработаны экспериментальные стратегии производить антитела, катализирующие химические реакции.Эти каталитические антитела или абзимы выбираются из моноклональных антител, генерируемых иммунизацией мышей гаптенами, которые имитируют переходные состояния реакций, катализируемых ферментами.
ПРИРОДНЫЕ АБЗИМЫ Первые природные абзимы были получены путем очистки антител из сыворотки крови человека. Антитела с протеазной активностью против вазоактивного кишечного пептида (VIP) были впервые выделены в сыворотке пациентов с астмаастмой. Было также показано, что эта активность VIP-азы присутствует в белках, которые представляют собой моноклональные легкие цепи человека, обнаруженные в моче пациентов с множественной миеломой.. Гидролизующие ДНК аутоантитела были также выделены из сывороток пациентов с системной красной волчанкой или ревматоидным артритом.
ПРИМЕНЕНИЕ • Большой потенциал в фармацевтической промышленности. • Детоксикация кокаина. • Специфическое нацеливание на уникальные детерминанты раковых клеток, называемые антигенами опухолевых клеток, на их поверхности, которые отсутствуют в нормальных клетках. • Одно приложение может касаться использования гидролитических свойств абзимов для активации пролекарств.Направляя эту активность вблизи опухолевых клеток, пролекарства могут быть преобразованы в цитотоксические соединения непосредственно на опухолевых клетках. Эта противораковая терапия разработана как терапия антителами, направленная на абзим-пролекарство (ADAPT). • Деактивировать вирусы. • Наконец, все исследования, направленные на получение антител с протеазной активностью, специфичной для последовательности, могут открыть новые пути для антивирусной терапии и создания новых вакцин.
СТРУКТУРА АТФ
- Ресурс исследования
- Исследовать
- Искусство и гуманитарные науки
- Бизнес
- Инженерная технология
- Иностранный язык
- История
- Математика
- Наука
- Социальная наука
Лучшие подкатегории
- Высшая математика
- Алгебра
- Основы математики
- Исчисление
- Геометрия
- Линейная алгебра
- Предалгебра
- Предварительный расчет
- Статистика и вероятность
- Тригонометрия
- другое →
Лучшие подкатегории
- Астрономия
- Астрофизика
- Биология
- Химия
- Науки о Земле
- Наука об окружающей среде
- Наука о здоровье
- Физика
- другое →
Лучшие подкатегории
- Антропология
- Закон
- Политология
- Психология
- Социология
- другое →
Microsoft Defender for Identity Security alert tutorial
- 5 минут на чтение
В этой статье
Важно
Названия продуктов защиты от угроз от Microsoft меняются.Подробнее об этом и других обновлениях читайте здесь. В ближайшем будущем мы обновим названия продуктов и документов.
Microsoft Defender for Identity предупреждает с помощью понятного языка и графики, какие подозрительные действия были выявлены в вашей сети, а также участники и компьютеры, вовлеченные в угрозы. Оповещения классифицируются по степени серьезности, имеют цветовую кодировку, чтобы их было легко визуально фильтровать, и упорядочены по фазам угроз. Каждое предупреждение разработано, чтобы помочь вам быстро понять, что происходит в вашей сети.Списки свидетельств оповещения содержат прямые ссылки на вовлеченных пользователей и компьютеры, чтобы облегчить и упростить ваши расследования.
В этом руководстве вы узнаете о структуре предупреждений безопасности Defender for Identity и о том, как их использовать:
- Структура оповещения системы безопасности
- Классификация предупреждений системы безопасности
- Категории предупреждений системы безопасности
- Исследование Advanced Security Alert
- Связанные предприятия
- Защитник для идентификации и NNR (разрешение сетевого имени)
Структура оповещения безопасности
Каждое оповещение безопасности Defender for Identity включает:
- Alert title
Official Defender for Identity имя предупреждения. - Описание
Краткое объяснение произошедшего. - Доказательства
Дополнительная соответствующая информация и соответствующие данные о том, что произошло, чтобы помочь в процессе расследования. - Excel скачать
Подробный отчет Excel скачать для анализа
Классификация предупреждений безопасности
После надлежащего расследования все предупреждения системы безопасности Defender for Identity можно отнести к одному из следующих типов активности:
Истинно положительное (TP) : вредоносное действие, обнаруженное Защитником идентификации.
Доброкачественный истинно положительный результат (B-TP) : Действие, обнаруженное Defender for Identity, которое является реальным, но не вредоносным, например тест на проникновение или известная активность, созданная утвержденным приложением.
Ложное срабатывание (FP) : Ложная тревога, означающая, что действия не произошло.
Является ли предупреждение системы безопасности TP, B-TP или FP
Для каждого предупреждения задайте следующие вопросы, чтобы определить классификацию предупреждений и помочь решить, что делать дальше:
- Насколько распространено это конкретное предупреждение системы безопасности в вашей среде?
- Было ли предупреждение инициировано компьютерами или пользователями одного типа? Например, серверы с одинаковой ролью или пользователи из одной группы / отдела? Если компьютеры или пользователи были похожи, вы можете решить исключить это, чтобы избежать дополнительных предупреждений FP в будущем.
Примечание. Увеличение количества предупреждений одного и того же типа обычно снижает уровень подозрительности / важности предупреждения. Для повторяющихся предупреждений проверьте конфигурации и используйте сведения и определения предупреждений безопасности, чтобы точно понять, что происходит, вызывая повторение.
Категории предупреждений безопасности
Предупреждения системы безопасностиDefender for Identity делятся на следующие категории или фазы, как и фазы, наблюдаемые в типичной цепочке уничтожения кибератаки. Узнайте больше о каждом этапе и предупреждениях, предназначенных для обнаружения каждой атаки, используя следующие ссылки:
Расширенное расследование предупреждений системы безопасности
Чтобы получить более подробную информацию о предупреждении системы безопасности, загрузите подробный отчет по предупреждению в Excel.
- Щелкните три точки в правом верхнем углу любого предупреждения, выберите Подробнее о загрузке .
Каждая загрузка Excel для оповещения Defender for Identity предоставляет следующую информацию:
- Сводка - первая вкладка включает основные моменты предупреждения
- Название
- Описание
- Время начала (UTC)
- Время окончания (UTC)
- Уровень серьезности - низкий / средний / высокий
- Статус - открыто / закрыто
- Время обновления статуса (UTC)
- Просмотреть в браузере
- Все задействованные объекты (учетные записи, компьютеры и ресурсы) перечислены, разделенные по их ролям.
- Источник, место назначения или атакованный, в зависимости от предупреждения.
- Большинство вкладок содержат следующие данные по каждой сущности:
- Имя
- Детали
- Тип
- SamName
- Исходный компьютер
- Пользователь-источник (при наличии)
- Контроллеры домена
- Доступный ресурс: время, компьютер, имя, сведения, тип, услуга.
- Дополнительные вкладки для каждого предупреждения:
- На атакованные аккаунты, когда предполагаемая атака использовала грубую силу.
- На серверах системы доменных имен (DNS), когда подозреваемый атаковал, задействовал рекогносцировку сетевого сопоставления (DNS).
- Связанные объекты: ID, тип, имя, уникальный объект Json, уникальный профиль объекта Json
- Все необработанные действия, зафиксированные Defender for Identity Sensors, связанные с предупреждением (действия сети или события), включая:
- Сетевая деятельность
- События
Связанные предприятия
В каждом предупреждении последняя вкладка содержит Связанные объекты .Связанные объекты - это все объекты, вовлеченные в подозрительную деятельность, без разделения «роли», которую они сыграли в предупреждении. У каждого объекта есть два файла Json: уникальный объект Json и уникальный профиль объекта Json. Используйте эти два файла Json, чтобы узнать больше об объекте и помочь вам изучить предупреждение.
Уникальная сущность Json
Включает данные Defender for Identity, полученные из Active Directory об учетной записи. Сюда входят все атрибуты, такие как Distinguished Name , SID , LockoutTime и PasswordExpiryTime .Для учетных записей пользователей включает такие данные, как Department , Mail и PhoneNumber . Для учетных записей компьютеров включает такие данные, как OperatingSystem , IsDomainController и DnsName .
Уникальный профиль объекта Json
Включает все данные Defender for Identity, профилированные для объекта. Defender for Identity использует записанные сетевые операции и события, чтобы узнать о пользователях и компьютерах среды.Defender for Identity предоставляет соответствующую информацию для каждой организации. Эта информация помогает Defender for Identity определять угрозы.
Как я могу использовать Защитник для идентификации личности в расследовании?
Расследования могут быть сколь угодно подробными. Вот несколько идей о способах исследования с использованием данных, предоставленных Defender for Identity.
- Проверить, принадлежат ли все связанные пользователи к одной группе или отделу?
- Связанные пользователи совместно используют ресурсы, приложения или компьютеры?
- Активна ли учетная запись, хотя ее PasswordExpiryTime уже прошел?
Защитник для идентификации и NNR (разрешение сетевых имен)
ВозможностиDefender для определения личности полагаются на активное разрешение сетевых имен (NNR) для разрешения IP-адресов компьютерам в вашей организации.Используя NNR, Defender for Identity может сопоставлять необработанные действия (содержащие IP-адреса) и соответствующие компьютеры, участвующие в каждом действии. На основе необработанных действий Defender for Identity профилирует сущности, включая компьютеры, и генерирует предупреждения.
ДанныеNNR имеют решающее значение для обнаружения следующих предупреждений:
- Предполагаемая кража личных данных (проездной)
- Предполагаемая атака DCSync (репликация служб каталогов)
- Разведка сетевых карт (DNS)
Используйте информацию NNR, представленную на вкладке Network Activities отчета о загрузке предупреждений, чтобы определить, является ли предупреждение FP .В случае предупреждения FP обычно результат достоверности NNR предоставляется с низкой достоверностью.
Данные отчета о загрузке отображаются в двух столбцах:
Дополнительные сведения о работе с предупреждениями безопасности Defender for Identity см.
