Система сахаропосредованной регуляции и роль киназ HXK1, SnRK1, TOR у Arabidopsis thaliana | Бельков
1. Smeekens S., Ma J., Hanson J., Rolland F. Sugar signals and molecular networks controlling plant growth // Current Opinion in Plant Biology. 2010. Vol. 13. Issue 3. P. 274-279. https://doi.org/10.1016/j.pbi.2009.12.002
2. Matsoukas I.G. Interplay between sugar and hormone signaling pathways modulate floral signal transduction // Frontiers in Genetics. 2014. Vol. 5. P. 218. https://doi.org/10.3389/fgene.2014.00218
3. Moore B., Zhou L., Rolland F., Hall Q., Cheng W.-H., Liu Y.-X., et al. Role of the Arabidopsis glucose sensor HXK1 in nutrient, light, and hormonal signaling // Science. 2003. Vol. 300. Issue 5617. P. 332-336. https://doi.org/10.1126/science.1080585
4. Rolland F., Baena-Gonzalez E., Sheen J. Sugar sensing and signaling in plants: conserved and novel mechanisms // Annual Review of Plant Biology. 2006. Vol. 57. P. 675-709. https://doi.org/10.1146/annurev. arplant.57.032905.105441
arplant.57.032905.105441
5. Van Dijken A.J., Schluepmann H., Smeekens S.C. Arabidopsis trehalose-6-phosphate synthase 1 is essential for normal vegetative growth and transition to flowering // Plant Physiology. 2004. Vol. 135. Issue 2. P. 969-977. https://doi.org/10.1104/pp.104.039743
6. Baena-Gonzalez E., Rolland F., Thevelein J.M., Sheen J. A central integrator of transcription networks in plant stress and energy signaling // Nature. 2007. Vol. 448. Issue 7156. P. 938-942. https://doi.org/10.1038/nature06069
7. Schluepmann H., van Dijken A., Aghdasi M., Wobbes B., Paul M., Smeekens S. Trehalose mediated growth inhibition of Arabidopsis seedlings is due to trehalose-6-phosphate accumulation // Plant Physiology. 2004. Vol. 135. Issue 2. P. 879-890. https://doi.org/10.1104/pp.104.039503
8. Hausler R.E., Heinrichs L., Schmitz J., Flugge U.-I. How sugars might coordinate chloroplast and nuclear gene expression during acclimation to high light intensities // Molecular Plant. 2014. Vol. 7. Issue 7. P. 1121-1137. https://doi.org/10.1093/mp/ssu064
2014. Vol. 7. Issue 7. P. 1121-1137. https://doi.org/10.1093/mp/ssu064
9. Ramon M., Rolland F., Sheen J. Sugar sensing and signaling // The Arabidopsis Book. 2008. Vol. 6. P. e0117. https://doi.org/10.1199/tab.0117
10. Biasing O.E., Gibon Y., Gunther M., Hohne M., Morcuende R., Osuna D., et al. Sugars and circadian regulation make major contributions to the global regulation of diurnal gene expression in Ara-bidopsis // The Plant Cell. 2005. Vol. 17. Issue 12. P. 3257-3281. https://doi.org/10.1105/tpc.105.035261
11. Sami F., Siddiqui H., Hayat S. Interaction of glucose and phytochormone signaling in plants // Plant Physiology and Biochemistry. 2019. Vol. 135. P. 119-126. https://doi.org/10.1016/j.plaphy.2018.11.005
12. Xiong Y., McCormack M., Li L., Hall Q., Xiang C., Sheen J. Glc-TOR signaling leads transcriptome reprogramming and meristem activation // Nature. 2013. Vol. 496. Issue 7444. P. 181-186. https://doi.org/10.1038/nature12030
13. Aguilera-Alvarado G.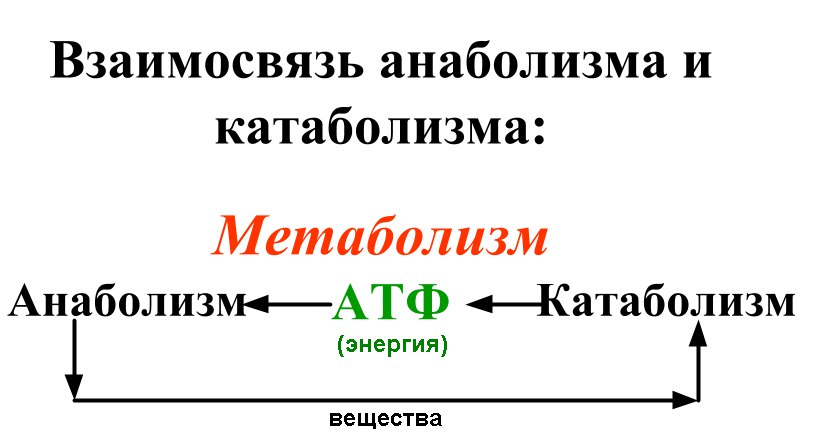 P., Sanchez-Nieto S. Plant hexokinases are multifaceted proteins // Plant and Cell Physiology. 2017. Vol. 58. Issue 7. P. 11511160. https://doi.org/10.1093/pcp/pcx062
P., Sanchez-Nieto S. Plant hexokinases are multifaceted proteins // Plant and Cell Physiology. 2017. Vol. 58. Issue 7. P. 11511160. https://doi.org/10.1093/pcp/pcx062
14. Cho J.-I., Ryoo N., Eom J.S., Lee D.-W., Kim H.-B., Jeong S.-W., et al. Role of the rice hexokina-ses OsHXK5 and OsHXK6 as glucose sensors // Plant Physiology. 2009. Vol. 149. P. 745-759. https://doi.org/10.1104/pp.108.131227
15. Zhang C., Han L., Slewinski T.L., Sun J., Zhang J., Wang Z.-Y., et al. Symplastic phloem loading in poplar // Plant Physiology. 2014. Vol. 166. Issue 1. P. 306-313. https://doi.org/10.1104/pp.114.245845
16. Sheen J. Metabolic repression of transcription in higher plants // The Plant Cell. 1990. Vol. 2. P. 1027-1038. https://doi.org/10.1105/tpc.2.10.1027
17. Cho Y.-H., Yoo S.-D., Sheen J. Glucose signaling through nuclear hexokinase1 complex in Ara-bidopsis // Plant Signaling and Behavior. 2007. Vol. 2. Issue 2. P. 123-124. https://doi.org/10.1016/j.cell.2006.09.028
18. Rottmann T., Fritz C., Sauer N., Stadler R. Glucose uptake via STP transporters inhibits in vitro pollen tube growth in a HEXOKINASE1-dependent manner in Arabidopsis thaliana // The Plant Gell. 2018. Vol. 30. Issue 9. P. 2057-2081. https://doi.org/10.1105/tpc.18.00356
Rottmann T., Fritz C., Sauer N., Stadler R. Glucose uptake via STP transporters inhibits in vitro pollen tube growth in a HEXOKINASE1-dependent manner in Arabidopsis thaliana // The Plant Gell. 2018. Vol. 30. Issue 9. P. 2057-2081. https://doi.org/10.1105/tpc.18.00356
19. Kim J. Sugar metabolism as input signals and fuel for leaf senescence // Genes and Genomics. 2019. Vol. 41. Issue 7. P. 737-746. https://doi.org/10.1007/s13258-019-00804-y
20. Miao H., Cai C., Wei J., Chang J., Qian H., Zhang X., et al. Glucose enhances indolic glucosin-olate biosynthesis without reducing primary sulfur assimilation // Scientific Reports. 2016. Vol. 6. Article no. 31854. https://doi.org/10.1038/srep31854
21. Huang W., Yu C., Hu J., Wang L., Dan Z., Zhou W., et al. Pentatricopeptide-repeat family protein RF6 functions with hexokinase 6 to rescue rice cytoplasmic male sterility // Proceedings of the National Academy of Sciences of the United States of America. 2015. Vol. 112. P. 14984-14989. https://doi.org/10.1073/pnas.1511748112
https://doi.org/10.1073/pnas.1511748112
22. Rook F., Handingham S.A., Li Y., Bevan M.W. Sugar and ABA response pathways and the control of gene expression // Plant Cell & Environment. 2006. Vol. 29. Issue 3. P. 426-434. https://doi.org/10.1111/j.1365-3040.2005.01477.x
23. Arenas-Huertero F., Arroyo A., Zhou L., Sheen J., Leon P. Analysis of Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar // Genes and Development. 2000. Vol. 14. Issue 16. P. 2085-2096.
24. Koussevitzky S., Nott A., Mockler T.C., Hong F., Sachetto-Martins G., Surpin M., et al. Signals from chloroplasts converge to regulate nuclear gene expression // Science. 2007. Vol. 316. Issue 5825. P. 715-719. https://doi.org/10.1126/science.1140516
25. Gregorio J., Hernandez-Bernal A.F., Cordoba E., Leon P. Characterization of evolutionarily conserved motifs involved in activity and regulation of the ABA-INSENSITIVE (ABI) 4 transcription factor // Molecular Plant. 2014. Vol. 7. Issue 2. P. 422-436. https://doi.org/10.1093/mp/sst132
2014. Vol. 7. Issue 2. P. 422-436. https://doi.org/10.1093/mp/sst132
26. Giraud E., van Aken O., Ho L.H.M., Whelan J. The transcription factor ABI4 is a regulator of mitochondrial retrograde expression of ALTERNATIVE OXIDASE1a // Plant Physiology. 2009. Vol. 150. Issue 3. P. 1286-1296. https://doi.org/10.1104/pp.109. 139782
27. Leon P., Gregorio J., Cordoba E. ABI4 and its role in chloroplast retrograde communication // Frontiers in Plant Science. 2013. Vol. 3. Article 304. 13 p. https://doi.org/10.3389/fpls.2012.00304
28. Tsai A.Y.-L., Gazzarrini S. Trehalose-6-phosphate and SnRK1 kinases in plant development and signaling: the emerging picture // Frontiers in Plant Science. 2014. Vol. 5. Article 119. 11 p. https://doi.org/10.3389/fpls.2014.00119
29. Swartzberg D., Hanael R., Granot D. Rela tionship between hexokinase and cytokinin in the regulation of leaf senescence and seed germination // Plant Biology. 2011. Vol. 13. Issue 3. P. 439-444. https://doi.org/10.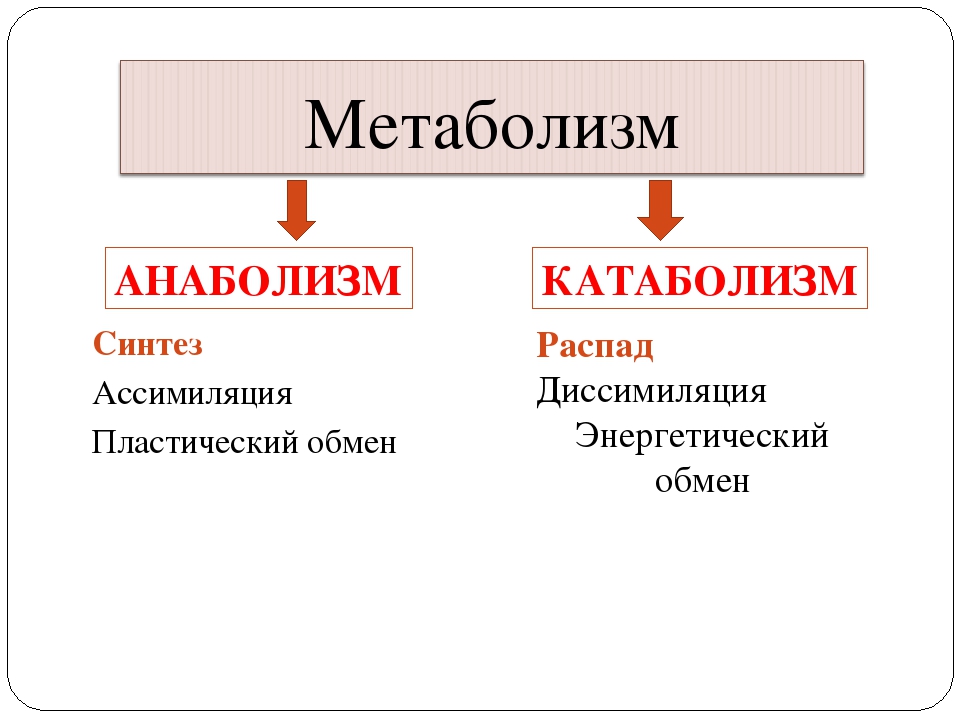 1111/j.1438-8677.2010.00376.x
1111/j.1438-8677.2010.00376.x
30. Li L., Sheen J. Dynamic and diverse sugar signaling // Current Opinion in Plant Biology. 2016. Vol. 33. P. 116-125. https://doi.org/10.1016/j.pbi. 2016.06.018
31. Crozet P., Margalha L., Confraria A., Rodrigues A., Martinho C., Adamo M., et al. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases // Frontiers in Plant Science. 2014. Vol. 5. P. 190. https://doi.org/10.3389/fpls.2014.00190
32. Glab N., Qury C., Guerinier T., Domenichini S., Crozet P., Thomas M., et al. The impact of Ara-bidopsis thaliana SNF1-related-kinase 1 (SnRK1)-activating kinase 1 (SnAK1) and SnAK2 on SnRK1 phosphorylation status: characterization of a SnAK double mutant // The Plant Journal. 2017. Vol. 89. Issue 5. P. 1031-1041. https://doi.org/10.1111/tpj.13445
33. Blanco N.E., Liebsch D., Diaz M.G., Strand A., Whelan J. Dual and dynamic intracellular localization of Arabidopsis thaliana SnRK1.1 // Journal of Experimental Botany. 2019. Vol. 70. Issue 8. P. 2325-2338. https://doi.org/10.1093/jxb/erz023
P. 2325-2338. https://doi.org/10.1093/jxb/erz023
34. Van Dingenen J., Vermeersch M., de Milde L., Hulsmans S., de Winne N., van Leene J., et al. The role of HEXOKINASE1 in Arabidopsis leaf growth // Plant Molecular Biology. 2019. Vol. 99. Issue 1-2. P. 79-93. https://doi.org/10.1007/s11103-018-0803-0
35. O’Hara L.E., Paul M.J., Wingler A. How do sugars regulate plant growth and development? New insight into the role of trehalose-6-phosphate // Molecular Plant. 2013. Vol. 6. Issue 2. P. 261-274. https://doi.org/10.1093/mp/sss120
36. Dong P., Xiong F., Que Y., Wang K., Yu L., Li Z., et al. Expression profiling and functional analysis reveals that TOR is a key player in regulating photosynthesis and phytohormone signaling pathways in Arabidopsis // Frontiers in Plant Science. 2015. Vol. 6. Article 667. 15 p. https://doi.org/10.3389/fpls.2015.00677
37. Dobrenel T., Caldana C., Hanson J., Robaglia C., Vincentz M., Viet B., et al. TOR signaling and nutrient sensing // Annual Review of Plant Biology. 2016. Vol. 67. P. 261-285. https://doi.org/10.1146/annurev-arplant-043014-114648
2016. Vol. 67. P. 261-285. https://doi.org/10.1146/annurev-arplant-043014-114648
38. Cao P., Kim S.-J., Xing A., Schenck C.A., Liu L., Jiang N., et al. Homeostasis of branched-chain amino acids is critical for the activity of TOR signaling in Arabidopsis // eLife. 2019. Vol. 8. P. e50747. https://doi.org/10.7554/eLife.50747
39. Shinkawa H., Kajikawa M., Nomura Y., Ogura M., Sawaragi Y., Yamano T., et al. Algal protein kinase, triacylglycerol accumulation regulator1, modulates cell viability and gametogenesis in car-bon/nitrogen imbalanced conditions // Plant Cell Physiology. 2019. Vol. 60. Issue 4. P. 916-930. https://doi.org/10.1093/pcp/pcz010
40. Paul M.J., Jhurreea D., Zhang Y., Primave-si L.F., Delatte T., Schluepmann H., et al. Upregulation of biosynthetic processes associated with growth by trehalose 6-phosphate // Plant Signaling & Behavior. 2010. Vol. 5. Issue 4. P. 386-392. https://doi.org/10.4161/psb.5.4.10792
41. Delatte T.L., Sedijani P., Kondou Y. , Matsui M., de Jon G.J., Somsen G.W., et al. Growth arrest by trehalose-6-phosphate: an astonishing case of primary metabolite control over growth by way of the SnRK1 signaling pathway // Plant Physiology. 2011. Vol. 157. Issue 1. P. 160-174. https://doi.org/10.1104/pp.111.180422
, Matsui M., de Jon G.J., Somsen G.W., et al. Growth arrest by trehalose-6-phosphate: an astonishing case of primary metabolite control over growth by way of the SnRK1 signaling pathway // Plant Physiology. 2011. Vol. 157. Issue 1. P. 160-174. https://doi.org/10.1104/pp.111.180422
42. Yadav U.P., Ivakov A., Feil R., Duan G.Y., Walther D., Giavalisco P., et al. The sucrosetrehalose 6-phosphate (Tre6P) nexus: specificity and mechanisms of sucrose signalling by Tre6P // Journal of Experimental Botany. 2014. Vol. 65. Issue 4. P. 1051-1068. https://doi.org/10.1093/jxb/ert457
43. Cho Y.-H., Hong J.-W., Kim E.-C., Yoo S.-D. Regulatory functions of SnRK1 in stress-responsive gene expression and in plant growth and development // Plant Physiology. 2012. Vol. 158. Issue 4. P. 1955-1964. https://doi.org/10.1104/pp.111.189829
44. Moreau M., Azzopardi M., Clement G., Dobrenel T., Marchive C., Renne C., et al. Mutations in the Arabidopsis homolog of LST8/GeL, a partner of the target of rapamycin kinase, impair plant growth, flowering, and metabolic adaptation to long days // The Plant Cell. 2012. Vol. 24. Issue 2. P. 463-481. https://doi.org/10.1105/tpc.111.091306
2012. Vol. 24. Issue 2. P. 463-481. https://doi.org/10.1105/tpc.111.091306
45. Xiong Y., McCormack M., Li L., Hall Q., Xiang C., Sheen J. Glucose-TOR signalling reprograms the transcriptome and activates meristems // Nature. 2013. Vol. 496. Issue7444. P. 181-186. https://doi.org/10.1038/nature12030
46. Baena-Gonzalez E., Rolland F., Thevelein J.M., Sheen J. A central integrator of transcription networks in plant stress and energy signaling // Nature. 2007. Vol. 448. Issue 7156. P. 938-942. https://doi.org/10.1038/nature06069
47. Karve A., Xia X., Moore B. Arabidopsis Hexokinase-Like1 and Hexokinase1 form a critical node in mediating plant glucose and ethylene responses // Plant Physiology. 2012. Vol. 158. P. 1965-1975. https://doi.org/10.1104/pp.112.195636
48. Kelly G., David-Schwartz R., Sade N., Moshelion M., Levi A., Alchanatis V., et al. The Pitfalls of Transgenic Selection and New Roles of AtHXK1: A high level of AtHXK1 expression uncouples hexokinase1-dependent sugar signaling from exogenous sugar // Plant Physiology. 2012. Vol. 159. Issue 1. P. 47-51. https://doi.org/10.1104/pp.112.196105
2012. Vol. 159. Issue 1. P. 47-51. https://doi.org/10.1104/pp.112.196105
49. Wingler A., Delatte T.L., O’Hara L.E., Primavesi L.F., Jhurreea D., Paul M.J., et al. Trehalose 6-phosphate is required for the onset of leaf senescence associated with high carbon availability // Plant Physiology. 2012. Vol. 158. P. 1241-1251. https: //doi.org/10.1104/pp.111.191908
50. Salem M.A., Li Y., Wiszniewski A., Giavalisco P. Regulatory-associated protein of TOR (RAPTOR) alters the hormonal and metabolic composition of Arabidopsis seeds, controlling seed morphology, viability and germination potential // The Plant Journal. 2017. Vol. 92. Issue 4. P. 525-545. https://doi.org/10.1111/tpj.13667
?Рыбий жир и анаболизм!
Как правило, пищевые добавки используются для достижения какого-то конкретного результата, например, для снижения веса, наращивания мышц или для общего оздоровления организма. Однако, существуют и исключения. К таким исключениям относится рыбий жир.
Всем известно благотворное влияние рыбьего жира на здоровье сердечно-сосудистой системы. Полезен рыбий жир и в борьбе с лишним весом. В этой статье мы рассмотрим потенциальное применение рыбьего жира для увеличения мышечной массы.
Антикатаболическое действие рыбьего жира
Белок мышц подвергается бесконечному процессу синтеза (анаболизма) и разрушения (катаболизма). В здоровом состоянии анаболический и катаболический процессы сбалансированы для поддержания определенного количества мышечной массы или даже ее увеличения (что наблюдается при сочетании тренинга с отягощениями и правильного питания).
Катаболизм мышечной ткани является обычным явлением при различных клинических состояниях (диабет, почечная недостаточность, травмы, рак), а также во время потери веса в результате диеты и в других стрессовых состояниях. В таких катаболических состояниях процесс разрушения мышечного белка опережает процесс его синтеза, что ведет к потере мышечной массы и истощению.
В первую очередь катаболизм мышечного протеина вызывается убиквитин-протеасомной системой. И тут на помощь может прийти рыбий жир, так как содержащаяся в нем эйкозапентаеновая кислота (ЭПК) значительно снижает активность катаболической (убиквитин-протеасомной) системы, разрушающей мышечный белок.
Другой механизм, при помощи которого рыбий жир оказывает своё антикатаболическое действие, это снижение уровня кортизола. Как все мы знаем, кортизол расщепляет мышечную ткань. Кроме того, постоянно высокий уровень кортизола оказывает негативное воздействие на организм, поэтому благотворное воздействие рыбьего жира не ограничивается только антикатаболическим эффектом.
Анаболическое действие рыбьего жира
Рыбий жир интересен ещё и тем, что способствует росту мышц не только посредством подавления мышечного катаболизма. Рыбий жир также стимулирует мышечный анаболизм. Недавние исследования показали, что, принимая в течение 8 недель ежедневно по 4 г концентрата рыбьего жира, обеспечивающего дневную дозу 1,86 г ЭПК (эйкозапентаеновой кислоты) и 1. 5 г ДГК (докозагексаеновой кислоты), можно значительно увеличить анаболическую реакцию синтеза мышечного белка. Доказано, что усиление анаболической реакции на аминокислоты и инсулин происходит за счёт повышения активации сигнального пути mTOR/p70S6K, который имеет важное значение для анаболизма мышечного белка и роста мышечных клеток.
5 г ДГК (докозагексаеновой кислоты), можно значительно увеличить анаболическую реакцию синтеза мышечного белка. Доказано, что усиление анаболической реакции на аминокислоты и инсулин происходит за счёт повышения активации сигнального пути mTOR/p70S6K, который имеет важное значение для анаболизма мышечного белка и роста мышечных клеток.
Вероятно, задействованы и другие механизмы. В том же исследовании доказывается, что приём рыбьего жира здоровыми людьми в возрасте от 24 до 45 лет вдвое увеличивает соотношение ЭПК, ДПК (ещё одна, менее известная, жирная омега-3 кислота) и ДГК в мембранах мышечных клеток за счёт омега-6 кислот и мононенасыщенных жирных кислот. Концентрация насыщенных жирных кислот при этом не изменяется. Таким образом, вполне вероятно, что включение в рацион питания рыбьего жира оказывает влияние на анаболические сигнальные каскады, воздействуя на липидный состав и/или подвижность мембран.
Вам уже за 45? Не волнуйтесь, вам тоже пойдёт на пользу анаболическое воздействие рыбьего жира на мышцы. Тот же исследовательский коллектив провёл такой же эксперимент (1,86 г ЭПК и 1,5 г ДГК в течение 8 недель) с участием здоровых людей старше 65 лет (средний возраст испытуемых составил 71 год). Полученные результаты совпали с данными молодой возрастной группы. Включение в рацион рыбьего жира вызвало усиление анаболической реакции на аминокислоты и инсулин. Таким образом, рыбий жир, видимо, ослабляет сопротивление анаболизму, характерное для пожилого возраста. Результаты настолько поразили исследователей, что они сделали заключение: рыбий жир может быть полезен как при профилактике, так и при лечении саркопении (возрастное атрофическое дегенеративное изменение скелетной мускулатуры).
Тот же исследовательский коллектив провёл такой же эксперимент (1,86 г ЭПК и 1,5 г ДГК в течение 8 недель) с участием здоровых людей старше 65 лет (средний возраст испытуемых составил 71 год). Полученные результаты совпали с данными молодой возрастной группы. Включение в рацион рыбьего жира вызвало усиление анаболической реакции на аминокислоты и инсулин. Таким образом, рыбий жир, видимо, ослабляет сопротивление анаболизму, характерное для пожилого возраста. Результаты настолько поразили исследователей, что они сделали заключение: рыбий жир может быть полезен как при профилактике, так и при лечении саркопении (возрастное атрофическое дегенеративное изменение скелетной мускулатуры).
В обоих исследованиях количество мышечной массы не измерялось, так как эксперимент продолжался всего 8 недель. Однако, учитывая, что изменение метаболизма мышечного белка предшествует соответствующим изменениям мышечной массы, эти результаты обнадёживают. Будет интересно узнать о результатах более длительного исследования, в котором будут проведены вычисления фактической прибавки мышечной массы, вызванной приемом рыбьего жира. Кроме того, интересно узнать о взаимодействии анаболического отклика на прием рыбьего жира и тренировок с отягощениями.
Кроме того, интересно узнать о взаимодействии анаболического отклика на прием рыбьего жира и тренировок с отягощениями.
Факты свидетельствуют о необходимости включения в рацион рыбьего жира, независимо от того, хотите вы нарастить мышечную массу или предотвратить её потерю во время диеты. Рыбий жир и особенно ЭПК не только противодействуют потере мышечной массы в стрессовых и катаболических состояниях, но и повышают анаболическую реакцию на пищевые стимулы в здоровых мышцах, как у молодых людей, так и у лиц среднего и преклонного возраста. Таким образом, рыбий жир положительно влияет на баланс катаболических и анаболических процессов.
Справка по биохимии
Студенты, нуждающиеся в помощи по биохимии, получат большую пользу от нашей интерактивной программы. Мы разбираем все ключевые элементы, чтобы вы могли получить адекватную помощь по биохимии. Имея под рукой обязательные концепции обучения и актуальные практические вопросы, вы получите много помощи по биохимии в кратчайшие сроки. Получите помощь сегодня с нашей обширной коллекцией необходимой информации по биохимии.
Получите помощь сегодня с нашей обширной коллекцией необходимой информации по биохимии.Если вы в настоящее время изучаете биохимию или собираетесь изучать ее в будущем, учебные инструменты Varsity Tutors’ Learning Tools предоставляют ряд ресурсов, которые помогут вам в подготовке к биохимии. Планируете ли вы сдавать предметный тест по биохимии университетского уровня или просто экзамен для класса по биохимии, вы можете укрепить свои знания по этому предмету с помощью помощи по изучению биохимии, доступной для вас на веб-сайте средств обучения.
Некоторым студентам изучение биохимии может показаться трудным и непосильным. Вам нужно будет получить много информации о биохимии, клеточной биологии, молекулярной биологии и генетике. К счастью, у вас есть бесплатный доступ к интерактивной учебной программе по биохимии через ресурс Learn by Concept на веб-сайте Varsity Tutors’ Learning Tools. По мере того, как вы перемещаетесь по бесплатным учебным материалам по биохимии, предоставленным вам через ресурс Learn by Concept, вы заметите, что весь материал разбит на категории и подразделы.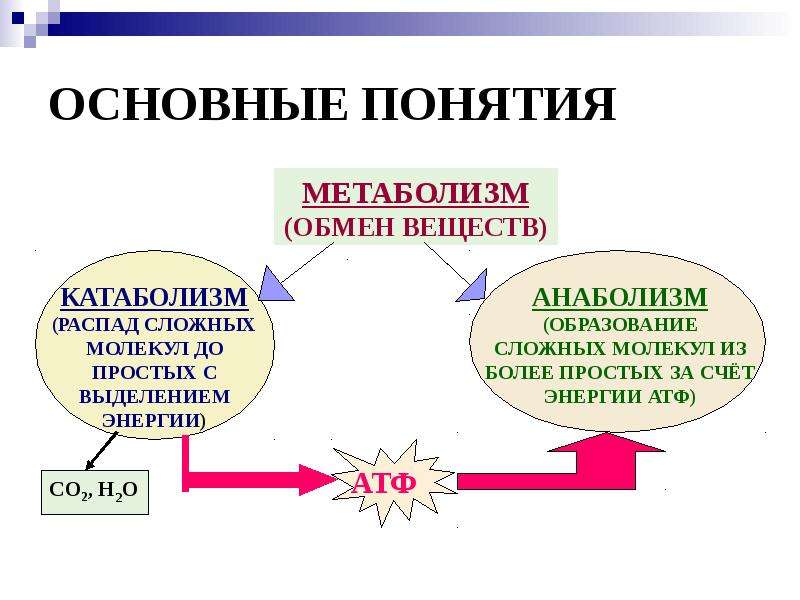 Вы можете просмотреть ряд тем, таких как анаболические и катаболические пути, биохимические передачи, метаболизм, кинетика ферментов и макромолекулы. Инструмент обучения «Узнай по концепции» очень всеобъемлющий и охватывает широкий круг тем, связанных с биохимией.
Вы можете просмотреть ряд тем, таких как анаболические и катаболические пути, биохимические передачи, метаболизм, кинетика ферментов и макромолекулы. Инструмент обучения «Узнай по концепции» очень всеобъемлющий и охватывает широкий круг тем, связанных с биохимией.
Learn by Concept не только задает вам вопросы по важному материалу, но и предоставляет важную информацию посредством подробных описаний. Эти описания помогают объяснить, почему предоставленный ответ является правильным, и помогают закрепить уже имеющиеся у вас знания. Используя помощь в изучении биохимии, вы можете точно определить области, над которыми вам нужно больше работать, и полностью настроить свой план изучения биохимии. Кроме того, вы можете узнать много важных формул, определений и словарных терминов, прочитав описания, приведенные в конце каждого примера вопроса по биохимии.
Кроме того, вы можете узнать много важных формул, определений и словарных терминов, прочитав описания, приведенные в конце каждого примера вопроса по биохимии.
Ресурс Learn by Concept упрощает подготовку к биохимии. Вам предоставляется информация о ряде понятий, необходимых для понимания биохимии, таких как эукариоты и прокариоты, преобразование энергии, структуры связей, белки и нейротрансмиттеры. Если вы хотите просмотреть тест или выучить новый материал, полное использование бесплатных учебных материалов по биохимии в средствах обучения Varsity Tutors — идеальный способ дополнить вашу курсовую работу.
Вы можете еще больше улучшить свою подготовку к биохимии, используя ресурс Learn by Concept вместе с остальными предоставленными вам средствами обучения. Если вы используете карточки, отвечаете на вопрос дня и оцениваете свои знания с помощью одного из многочисленных практических тестов по биохимии, вы помогаете создать индивидуальный, всесторонний режим обучения. Объедините все инструменты вместе, и вы сможете легко настроить свои учебные планы, отслеживать свои успехи и определять, какие области вам нужно уделить больше времени для улучшения.
Биохимия
Анаболические пути и синтез
Синтез углеводов
Анаболизм углеводов
Регулирование синтеза углеводов
Синтез липидов
Ферменты синтеза липидов
Реагенты, промежуточные продукты и продукты для синтеза липидов
Регуляция синтеза липидов
Синтез нуклеиновых кислот
Репликация ДНК
Регулирование репликации ДНК
Регулирование транскрипции
Транскрипция
Белковый синтез
Регуляция синтеза белка
Перевод
Биохимическая сигнализация
-
Гормоны и нейротрансмиттеры
Другие гормональные пути
Пути пептидных гормонов
Пути стероидных гормонов
Биохимия синапсов
Пути передачи сигнала
Путь белка G
Интегрины
Лиганд-управляемые ионные каналы
Вторые вестники
Толл-подобные рецепторы
Путь тирозинкиназы
Катаболические пути и метаболизм
Метаболизм углеводов
Альтернативные пути
Ферментация и анаэробное дыхание
Этаноловая ферментация
Молочнокислое брожение
Глюконеогенез
Энергетика глюконеогенеза
Ферменты глюконеогенеза
Регуляция глюконеогенеза
Другие концепции глюконеогенеза
- Реактивы и продукты глюконеогенеза
Гликогенолиз
Гликогенолиз Энергетика
Ферменты гликогенолиза
Регламент гликогенолиза
Другие концепции гликогенолиза
Реагенты и продукты гликогенолиза
Цикл лимонной кислоты
Промежуточные углеводы цикла лимонной кислоты
Цикл лимонной кислоты Энергетика
Ферменты цикла лимонной кислоты
Другие концепции цикла лимонной кислоты
Реагенты и продукты цикла производства лимонной кислоты
Транспорт электронов и окислительное фосфорилирование
АТФ-синтаза
Энергетика электрон-транспортной цепи
Белки и комплексы электрон-транспортной цепи
НАДН и ФАДх3
Другие концепции электрон-транспортной цепи
Другие концепции окислительного фосфорилирования
-
Окислительное фосфорилирование Энергетика
Гликолиз
Промежуточные углеводы гликолиза
Гликолиз Энергетика
Ферменты гликолиза
Регламент гликолиза
Другие концепции гликолиза
Реактивы и продукты гликолиза
Фотосинтез
Цикл Кальвина
Промежуточные углеводы цикла Кальвина
Другие концепции цикла Кальвина
Легкие реакции
Световая реакция Энергетика
Другие концепции световой реакции
Катаболизм липидов
Катаболизм липидов Энергетика
Ферменты катаболизма липидов
Промежуточные продукты катаболизма липидов
Регламент катаболизма липидов
Другие концепции катаболизма липидов
Реагенты и продукты катаболизма липидов
Катаболизм белков
Другие концепции белкового катаболизма
Ферменты белкового катаболизма
Промежуточные продукты катаболизма белков
Регламент катаболизма белков
Реагенты и продукты катаболизма белков
Кинетика ферментов и ингибирование
Ферментная кинетика и модели
Основы ферментативной кинетики
Уравнение Михаэлиса-Ментен
Графики Михаэлиса-Ментен
Другие принципы кинетики
Шаг ограничения скорости
Вмакс и
кмТипы торможения
Конкурентное торможение
Идентификация типа ингибирования
Смешанное ингибирование
Неконкурентное торможение
Неконкурентное торможение
Фундаментальные макромолекулы и концепции
Гомеостаз и биологическая среда
Биологическая энергетика
Основные свойства углерода
Основные свойства воды
Отрицательный отзыв
Другие гомеостатические состояния
Регламент pH
Термодинамическое регулирование
Основы макромолекул
Аминокислоты и белки
Липиды
Моносахариды и углеводы
Нуклеиновые кислоты: ДНК и РНК
Молекулярные связи и силы
Ковалентные связи
Водородные связи
Гидрофобные взаимодействия
Другие межмолекулярные силы
Идентификация биохимических молекул
Идентификация по функции
Определение конкретных функций углеводов
Идентификация специфических функций липидов
Идентификация конкретных функций нуклеиновых кислот
Идентификация специфических функций белка
Идентификация по структуре
Идентификация специфических структур углеводов
Идентификация специфических липидных структур
Идентификация конкретных структур нуклеиновых кислот
Идентификация специфических белковых структур
Идентификация мономеров и димеров
Идентификация аминокислот
Идентификация дисахаридов
Идентификация моносахаридов
Идентификация азотистых оснований и производных
Макромолекулярные структуры и функции
Структура и функции углеводов
Альдозы и кетозы
Эпимеры, хиральность и аномерный углерод
Гликозидные связи
Структурные углеводы
Структуры и функции липидов
Классификация липидов
Жирные кислоты
Глицеролипиды и триглицериды
Другие классы липидов
Фосфолипиды
Сфинголипиды
Стерины и стероиды
Структуры и функции нуклеиновых кислот
Деградация нуклеиновой кислоты
ДНКазы
Регуляция деградации нуклеиновых кислот
Структуры нуклеиновых кислот
ДНК и РНК
Модификации и эпигенетика
Азотистые основания
Производные нуклеиновой кислоты
Сахарофосфатные группы и фосфодиэфирные связи
Структура и функции белка
Деградация белка
Гидролиз белка
Регулирование деградации белка
Путь убиквитинирования
Свертывание белков
Первичная структура
Четвертичная структура
Регламент и шаперонины
Вторичная структура
Третичная структура
Белковые функции
Гидролазы
Изомеры
Лигазы
Другие функции белка
Структурные белки
Трансферазы и киназы
Транспортные белки
Белковая структура
Активный сайт
Глобулярные и волокнистые белки
Пептидные связи
Анаболический и катаболический (клеточный метаболизм): определение и примеры
Клетки — это мельчайшие единицы живых существ, обладающие всеми свойствами, присущими жизни. Одной из этих определяющих характеристик является метаболизм , или использование молекул или энергии, собранных из окружающей среды, для проведения биохимических реакций, необходимых для сохранения жизни и, в конечном счете, размножения.
Одной из этих определяющих характеристик является метаболизм , или использование молекул или энергии, собранных из окружающей среды, для проведения биохимических реакций, необходимых для сохранения жизни и, в конечном счете, размножения.
Метаболические процессы, часто называемые метаболическими путями, можно разделить на те, которые являются анаболическими или включают синтез новых молекул, и те, которые являются анаболическими0765 катаболические , которые включают расщепление существующих молекул.
Говоря простым языком, анаболические процессы связаны со строительством дома и заменой таких вещей, как окна и водосточные желоба по мере необходимости, а катаболические процессы связаны с устранением изношенных или сломанных частей дома. Если все это делать согласованно и в правильном темпе, дом будет существовать в максимально стабильном состоянии, но никогда не будет пассивным.
Обзор метаболизма
Клетки и ткани, которые они образуют, постоянно подвергаются «двунаправленному» метаболизму, а это означает, что в то время как одни вещи текут в анаболическом направлении, другие идут в противоположном направлении.
Это, возможно, более очевидно на уровне целых организмов: если вы сжигаете глюкозы во время бега, чтобы догнать свою собаку (катаболический процесс), бумага, порезанная на вашей руке накануне, продолжает заживать (анаболический процесс). Но та же самая дихотомия действует и в отдельных клетках.
Клеточные реакции катализируются особыми глобулярными белковыми молекулами, называемыми ферментами , которые по определению участвуют в химических реакциях, не изменяясь сами в конце. Они значительно ускоряют реакцию — иногда более чем в тысячу раз — и, таким образом, действуют как катализаторы .
Анаболические реакции обычно требуют затрат энергии и поэтому являются эндотермическими (в вольном переводе «тепло внутрь»). Это имеет смысл; вы не можете расти или наращивать мышечную массу, если не едите, при этом потребление пищи обычно масштабируется в зависимости от интенсивности и продолжительности данной деятельности.
Катаболические реакции обычно являются экзотермическими («тепло наружу») и высвобождают энергию, большая часть которой используется клеткой в форме аденозинтрифосфата (АТФ) и используется для других метаболических процессов.
Субстраты метаболизма
Основные структурные элементы тела и молекулы, необходимые ему для питания, а также для роста и замены тканей, состоят из мономеров или небольших повторяющихся единиц внутри большего целого, называемого полимером .
Эти единицы могут быть идентичными, как в случае с молекулами глюкозы, организованными в длинные цепочки запасного топлива гликогена , или они могут быть похожими и иметь «вкус», как в случае с нуклеиновыми кислотами и нуклеотидами, из которых они состоят.
Три основных класса макроэлементов макромолекул в питании человека, называемых углеводами , белками и жирами , каждый из которых состоит из мономера своего типа.
Глюкоза является основным субстратом всей жизни на Земле, и каждая живая клетка способна перерабатывать ее для получения энергии. Как уже отмечалось, молекулы глюкозы могут соединяться в «цепочки» с образованием гликогена, который у человека содержится преимущественно в мышцах и печени. Белки состоят из мономеров, полученных из набора из 20 различных аминокислот.
Жиры не являются полимерами, поскольку они состоят из трех жирных кислот, связанных с «основой» трехуглеродной молекулы глицерина . Когда они растут или сжимаются, это происходит за счет добавления или удаления атомов на концах цепочек жирных кислот, подобно заглавной букве «Е», когда вертикальная часть остается того же размера, а горизонтальные полосы различаются по длине.
Что такое анаболический метаболизм?
Подумайте о том, чтобы получить коробку игрушечных строительных блоков неограниченного размера. Многие идентичны, за исключением цвета; другие имеют разные размеры, но могут быть соединены вместе; третьи не предназначены для подключения независимо от выбранной вами конфигурации. Вы можете создавать идентичные конструкции, включающие, скажем, от трех до пяти частей, и связывать их вместе таким образом, чтобы соединения этих конструкций также были идентичными.
Вы можете создавать идентичные конструкции, включающие, скажем, от трех до пяти частей, и связывать их вместе таким образом, чтобы соединения этих конструкций также были идентичными.
По сути, это анаболический метаболизм в действии. Отдельные группы из трех-пяти фигурок представляют собой «мономеры», а готовый продукт аналогичен «полимеру». И в клетках, вместо того, чтобы ваши руки собирали кусочки вместе, ферменты руководят процессом. В обоих случаях ключевым аспектом является подвод энергии для создания молекул большей сложности (и, как правило, большего размера).
Примеры анаболических процессов включают, помимо синтеза белка, глюконеогенез (синтез глюкозы из различных вышестоящих субстратов), синтез жирных кислот, липогенез (синтез жиров из жирных кислот и глицерина) и образование мочевины и кетоновых тел .
Что такое катаболический метаболизм?
В большинстве случаев катаболические процессы на уровне индивидуальных реакций не являются просто соответствующими анаболическими реакциями, протекающими в обратном направлении, хотя многие из них совпадают. Обычно в процесс вовлечены различные ферменты.
Обычно в процесс вовлечены различные ферменты.
Например, первой стадией гликолиза (катаболизма глюкозы) является присоединение фосфатной группы к глюкозе с помощью фермента гексокиназы с образованием глюкозо-6-фосфата. Но последний этап глюконеогенеза, удаление фосфата из глюкозо-6-фосфата с образованием глюкозы, катализируется глюкозо-6-фосфатазой.
Другими жизненно важными катаболическими процессами, происходящими в вашем организме, являются гликогенолиз (распад гликогена в мышцах или печени), липолиз (удаление жирных кислот из глицерина), бета-окисление («сжигание» жирных кислот) и расщепление кетонов, белков или отдельных аминокислот.
Поддержание баланса анаболического и катаболического метаболизма
Поддержание тела в соответствии с его потребностями в режиме реального времени требует высокой степени реакции и координации. Скорости анаболических и катаболических реакций можно регулировать, изменяя количество фермента или субстрата, мобилизуемого в данную часть клетки, или посредством ингибирование по принципу обратной связи , при котором накопление продукта сигнализирует о более медленном протекании предшествующей реакции.
Кроме того, что важно с точки зрения целостной визуализации метаболизма, субстраты из одного пути макронутриентов могут быть переведены в другой по мере необходимости.
Примером такой интеграции путей является то, что аминокислоты аланин и глутамин, помимо того, что служат строительными блоками белков, также могут участвовать в глюконеогенезе. Для этого им нужно избавиться от азота, который обрабатывается ферментами, называемыми 9.0765 трансаминазы.
- Глицерин, продукт липолиза, также может участвовать в пути глюконеогенеза, который является одним из способов получения сахара из жира. Однако на сегодняшний день нет доказательств того, что продукты окисления жирных кислот могут участвовать в глюконеогенезе.
Физические упражнения: рост мышц и потеря жира
Физическая подготовка является серьезной общественной заботой в странах, где люди часто могут позволить себе роскошь дополнительных упражнений.
Многие из распространенных модальностей строго нацелены на тот или иной процесс, например, поднятие тяжестей для наращивания мышечной массы (анаболические упражнения) или использование эллиптического тренажера или беговой дорожки для «кардио» и сброс мышечной или жировой массы тела (или массы тела) для снижения веса (катаболические упражнения).
Одним из примеров работы обеих систем является подготовка марафонца к забегу на 42,2 км (26,2 мили). За неделю до этого многие люди намеренно наедались богатой углеводами пищей во время отдыха, чтобы потрудиться.
Из-за ежедневных беговых тренировок и постоянной потребности в восполнении катаболизированного топлива эти спортсмены имеют высокий уровень активности фермента гликогенсинтазы, что позволяет их мышцам и печени синтезировать гликоген с необычайной жадностью.
Во время марафона этот гликоген преобразуется в глюкозу, чтобы обеспечить бегуна часами подряд, хотя эти спортсмены обычно также принимают источники глюкозы (например, спортивные напитки) на протяжении всего забега, чтобы не «удариться о стену».
