Аминокислоты заменимые, незаменимые, условно заменимые, частично заменимые
Н. Разумовский Д. Соболев
Amino acids: nonessential and essential
N. Razumovsky D. Sobolev
Февраль 2020 • Корма
Скачать статью в PDF
Грамотное использование аминокислот в транзитный период способствует улучшению работы печени, укреплению иммунитета и поддержанию на оптимальном уровне обменных процессов, протекающих в организме высокопродуктивных коров.
Ключевые слова: аминокислотное питание высокопродуктивных коров в транзитный период, микробный белок, обменный белок, заменимые, частично заменимые, условно заменимые, незаменимые аминокислоты, расщепляемый и нерасщепляемый в рубце протеин, молочная продуктивность коров
Competent use of amino acids during the transit period contributes to liver function, improvement of immunity and maintaining optimum metabolic processes in the bodies of highly productive cows.
Кeywords: amino-acid nutrition of highly productive cows during transit period microbial protein metabolic protein nonessential partially nonessential conditionally nonessential essential amino acids rumen degradable protein and rumen nondegradable protein milk productivity of cows
2020-02-05
ИД «Животноводство»
[email protected]
Москва,
Хорошевское шоссе, 32 А, оф. 8
8
+7 901 578-71-29
8 800 551-73-54
«Обеспечиваем стабильный уровень жизни на селе»
Выращивание телят: кормление и профилактика диареи
Аналитические ИТ-инструменты для управления производством в мясной отрасли
Сохранность силоса и сенажа — 99%
Компания «Бионика» предлагает комплексное решение заготовки и хранения консервированных кормов — применение кислородно-барьерной полимерной пленки Силостоп производства компании Bruno Rimini.
Читать статью
Три линии защиты
Использование пробиотического препарата Alterion® компании Adisseo на основе B. subtilis (штамм DSM 29784) и защищенного бутирата позволяет уменьшить применение антибиотиков — стимуляторов роста и тем самым повысить эффективность сельхозпредприятий.
Читать статью
Вы готовы управлять своим будущим?
Использование технологии NeoTec4 и программы Nurture™ компании «Каргилл» позволяет выйти на новый уровень при выращивании молодняка крупного рогатого скота.
Читать статью
«Фидлэнд Групп» расширяет линейку экзогенных ферментов
ООО «Фидлэнд Групп» совместно с китайской биотехнологической корпорацией — одним из крупнейших мировых производителей ферментов VLAND BIOTECH GROUP — предлагает полный спектр ферментов, необходимых для производства сбалансированных комбикормов, отличающихся максимальной доступностью питательных
Читать статью
Нормализация функции кишечника
Применение кормовых добавок на основе богатой маннанами фракции клеточных стенок дрожжей позволяет оптимизировать состав микрофлоры ЖКТ за счет увеличения ее общего разнообразия, снижения патогенной нагрузки на кишечник и повышения его устойчивости к колонизации патогенами.
Читать статью
«Экономичность и эффективность — главные преимущества наших добавок»
Заместитель генерального директора по научно-исследовательской работе компании «Лаллеманд» (Россия) кандидат сельскохозяйственных наук Ирина Рябчик рассказывает об итогах работы компании на выставке «АГРОС-2022»: о полученной награде и организованном деловом семинаре.
Читать статью
Русская огранка — стандарт эталона
При использовании в кормлении коров бленда AMG-COMPLEX производства компании «АгроВитЭкс» увеличивается потребление СВ, повышается молочная продуктивность животных, укрепляется их иммунитет и улучшается воспроизводительная способность.
Читать статью
Осторожно: гербициды!
Ужесточение контроля за ввозом генно-модифицированной продукции в Россию позволит минимизировать вредное воздействие гербицидов на здоровье животных и человека.
Читать статью
Комплексный подход к кормлению коров
ООО «АгроВитЭкс» предлагает систему кормления коров, которая способствует повышению надоев, оздоровлению стада и помогает предотвратить болезни.
Читать статью
Наращиваем производство молока
Включение пробиотического препарата Профорт производства ООО «БИОТРОФ» в рационы для новотельных коров обеспечивает физиологически обоснованное течение метаболических процессов в их организме и способствует росту среднесуточных удоев и валового надоя молока натуральной жирности в первые три месяц
Читать статью
Гидролизат пера и мясо-костного остатка: источник белка для бройлеров
Ввод в рационы для бройлеров кормовой белковой добавки из кератинсодержащего сырья и использование пробиотического препарата Бацелл-М положительно сказались не только на продуктивности птицы, но и на ее мясных качествах.
Читать статью
Сохраняем питательность кормов с антиоксидантом Этоксол
Применение антиоксиданта Этоксол производства ГК «Апекс плюс» помогает защищать организм животных от действия свободных радикалов, поступления окисленных питательных веществ и повысить продуктивность.
Читать статью
Анимэйт®: контроль катионно-анионного баланса рационов в транзитный период
Компания Phibro Animal Health Corporation предлагает содержащую хлор и серу кормовую добавку Анимэйт® для высокопродуктивных коров, применение которой в транзитный период позволяет оптимизировать катионно-анионный баланс рациона и тем самым предотвратить развитие у животных гипокальциемии в субкл
Читать статью
Фитобиотики в кормлении животных
Фитобиотические препараты целесообразно использовать в животноводстве, в том числе в птицеводстве, в качестве альтернативы бактерицидным средствам.
Читать статью
Заготавливаем объемистые корма (продолжение)
Для создания высокопродуктивных пастбищ необходимо использовать бобово-злаковые травостои разных сроков созревания.
Читать статью
аргинин и гистидин. — Студопедия
Поделись с друзьями:
АМИНОКИСЛОТЫ
Белок организм напрямую не использует. Сначала белок расщепляется (гидролизуется) до аминокислот и аминокислотных групп (пептидов) и только затем эти «кирпичики» используются для восстановления/синтеза мышечных белков.
Аминокислоты — строительные блоки, из которых строятся мышечные волокна. Организм использует их для собственного роста, восстановления, укрепления и выработки различных гормонов, антител и ферментов.
Заменимые аминокислоты.

Заменимые аминокислоты: аланин, аргинин, аспарагин, аспарагиновая кислота, глицин (гликокол), глутамин, глутаминовая кислота, пролин, серин, тирозин, цистеин (цистин), цитруллин, гамма-аминомасляную кислоту, орнитин, таурин.
Частично заменимые аминокислоты: аргинин и гистидин.
Отличаются они от остальных тем, что организм может использовать их вместо, соответственно, метионина и фенилаланина для производства белка.
Существуют также аминокислоты, которые не синтезируются в организме человека, но необходимы для нормальной жизнедеятельности. Это — «незаменимые» аминокислоты.
Незаменимые аминокислоты: валин, лейцин, изолейцин, треонин, метионин, фенилаланин, триптофан, лизин
Они должны поступать в организм с пищей.Процесс синтеза белков постоянно идет в организме. В случае, когда хоть одна незаменимая аминокислота отсутствует, образование белков приостанавливается. Отсутствие или недостаток незаменимых аминокислот приводит к остановке роста, падению массы, нарушениям обмена веществ
Аминокислоты в свободной форме — это те аминокислоты, вы чаще всего видите на прилавках магазинов спортивного питания. .. Это аминокислоты, которые уже изначально переварены или расщеплены синтетическим путем. Однако вопреки заявлениям некоторых экспертов, потребление аминокислот в свободной форме — не лучший вариант получения протеина, необходимого для строительства новой мышечной ткани и поддержания тела в здоровом состоянии.
.. Это аминокислоты, которые уже изначально переварены или расщеплены синтетическим путем. Однако вопреки заявлениям некоторых экспертов, потребление аминокислот в свободной форме — не лучший вариант получения протеина, необходимого для строительства новой мышечной ткани и поддержания тела в здоровом состоянии.
Тем не менее, в некоторых обстоятельствах эти аминокислоты могут быть полезны, скажем, для достижения так называемых “специфических эффектов”. К примеру, некоторые аминокислоты, такие как триптофан и тирозин, оказывают прямое воздействие на нейротрансмиттеры. Потребление таких аминокислот в свободной форме, как глютамин и аргинин, способствуют повышению выработки гормона роста
Вам нужно понимать, что существует три вида аминокислотных добавок. BCAA, Комплексные и отдельные аминокислоты
ВСАА
Аббревиатура ВСАА означает Branched Chain Amino Acid, то есть, аминокислота с разветвленной боковой цепочкой. Эти аминокислоты являются самым важным компонентом белков мышц. Именно они препятствуют распаду мышечной ткани, вызванному интенсивным тренингом. ВСАА, а это три аминокислоты – изолейцин, лейцин и валин – являются незаменимыми аминокислотами, то есть, наш организм не способен их синтезировать, они могут поступать только извне. ВСАА могут использоваться мышцами и в качестве энергии, но такое их использование является крайне нецелесообразным. Именно поэтому совместно с приемом ВСАА нужно принимать достаточное количество простых углеводов.
Именно они препятствуют распаду мышечной ткани, вызванному интенсивным тренингом. ВСАА, а это три аминокислоты – изолейцин, лейцин и валин – являются незаменимыми аминокислотами, то есть, наш организм не способен их синтезировать, они могут поступать только извне. ВСАА могут использоваться мышцами и в качестве энергии, но такое их использование является крайне нецелесообразным. Именно поэтому совместно с приемом ВСАА нужно принимать достаточное количество простых углеводов.
Это продукты, содержащие в себе полный сбаллансированный набор аминокислот для построения мышечных белковых молекул. Подобные продукты включают в себя как заменимые, так и незаменимые аминокислоты в разных количествах.
Отдельные аминокислоты.
Не так распространены, как комплексные по причине своей более узкой специализации. Каждая отдельная аминокислота обладает определенным набором свойств. Давайте их рассмотрим.
Понравилась статья? Добавь ее в закладку (CTRL+D) и не забудь поделиться с друзьями:
Пищевая незаменимость «незаменимых аминокислот» для животных и человека
1. Wu G.
Функциональные аминокислоты для роста, размножения и здоровья. Ад Нутр
2010 г.; 1: 31–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Wu G.
Функциональные аминокислоты для роста, размножения и здоровья. Ад Нутр
2010 г.; 1: 31–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. Wu G. Аминокислоты: биохимия и питание, Бока-Ратон, Флорида: CRC Press, 2013. [Google Scholar]
3. Rose WC. Потребность в аминокислотах взрослого человека. Nutr Abstr Rev Ser Hum Exp 1957 год; 27: 631–47. [PubMed] [Академия Google]
4. Роуз туалет. Последовательность событий, ведущих к установлению потребности человека в аминокислотах. Am J Общественное здравоохранение 1968 год; 58: 2020–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Reeds PJ. Незаменимые и незаменимые аминокислоты для человека. Джей Нутр 2000 г.; 130: 1835–40 гг. [PubMed] [Google Scholar]
6. Бейкер Д.Х. Успехи в белково-аминокислотном питании птицы. Аминокислоты 2009 г.; 37: 29–41. [PubMed] [Google Scholar]
7. Абдерхальден Э.
Эксперимент по кормлению полностью разложившимися питательными веществами. Z Physiol Chem
1912; 77: 22–58. [Google Scholar]
[Google Scholar]
8. Роуз В.К., Кокс Г.Дж. Отношение аргинина и гистидина к росту. J Биол Хим 1924 г.; 61: 747–73. [Google Scholar]
9. Медицинский институт. Справочное потребление энергии, углеводов, клетчатки, жиров, жирных кислот, холестерина, белков и аминокислот, Вашингтон, округ Колумбия: The National Academies Press, 2005. [Google Scholar]
10. Morrison FB. Корма и кормление, 22-е изд. Итака, Нью-Йорк: The Morrison Publishing Company, 1956. [Google Scholar]
11. Maynard LA, Loosli JK, Hintz HF, Warner RG. Питание животных, 7-е изд. Нью-Йорк: McGraw-Hill Book Company, 1979. [Google Scholar]
12. Crampton EW, Harris LE. Прикладное питание животных, 2-е изд. Сан-Франциско: WH Freeman and Company, 1969. [Google Scholar]
13. Национальный исследовательский совет. Потребность свиней в питательных веществах. Вашингтон, округ Колумбия: National Academy Press, 2012. [Google Scholar]
14. Wu G, Wu ZL, Dai ZL, Yang Y, Wang WW, Liu C, Wang B, Wang JJ, Yin YL. Диетические потребности в «незаменимых аминокислотах» для животных и человека. Аминокислоты
2013; 44: 1107–13. [PubMed] [Академия Google]
Диетические потребности в «незаменимых аминокислотах» для животных и человека. Аминокислоты
2013; 44: 1107–13. [PubMed] [Академия Google]
15. Kalhan SC, Hanson RW. Возрождение серина: незаменимой аминокислоты, которой часто пренебрегают. J Биол Хим 2012 г.; 287: 19786–91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Phang JM, Liu W, Hancock C. Сближение эпигенетики и метаболизма: роль заменимых аминокислот. Эпигенетика 2013; 8: 231–236. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Wu G, Bazer FW, Dai ZL, Li DF, Wang JJ, Wu ZL. Аминокислотное питание животных: синтез белка и не только. Анну Рев Аним Биоски 2014; 2: 387–417. [PubMed] [Академия Google]
18. Li X, Bazer FW, Gao H, Jobgen W, Johnson GA, Li P, McKnight JR, Satterfield MC, Spencer TE, Wu G. Аминокислоты и газообразная передача сигналов. Аминокислоты 2009 г.; 37: 65–78. [PubMed] [Google Scholar]
19. Brosnan JT, Brosnan ME.
Креатин: эндогенный метаболит, диетическая и лечебная добавка. Анну Рев Нутр
2007 г.; 27: 241–61. [PubMed] [Google Scholar]
Анну Рев Нутр
2007 г.; 27: 241–61. [PubMed] [Google Scholar]
20. Даль Н., Пигг М., Ристофф Э., Гали Р., Карлссон Б., Маннервик Б., Ларссон А., Борд П. Миссенс-мутации в гене глутатионсинтетазы человека приводят к тяжелому метаболическому ацидозу, 5-оксопролинурии, гемолитической анемии и неврологической дисфункции. Хум Мол Жене 1997; 6: 1147–1152. [PubMed] [Google Scholar]
21. Ван Дж.Дж., Ву З.Л., Ли Д.Ф., Ли Н., Диндот С.В., Саттерфилд М.С., Базер Ф.В., Ву Г. Питание, эпигенетика и метаболический синдром. Антиоксидный окислительно-восстановительный сигнал 2012 г.; 17: 282–301. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Wang JJ, Chen LX, Li P, Li XL, Zhou HJ, Wang FL, Li DF, Yin YL, Wu G. Экспрессия генов изменяется в тонком кишечнике поросят при отъеме и добавлении в рацион глютамина. Джей Нутр 2008 г.; 138: 1025–1032. [PubMed] [Академия Google]
23. Джобген В., Фу В.Дж., Гао Х., Ли П., Майнингер К.Дж., Смит С.Б., Спенсер Т.Е., Ву Г.
Кормление с высоким содержанием жиров и добавка L-аргинина в пищу по-разному регулируют экспрессию генов в белой жировой ткани крыс.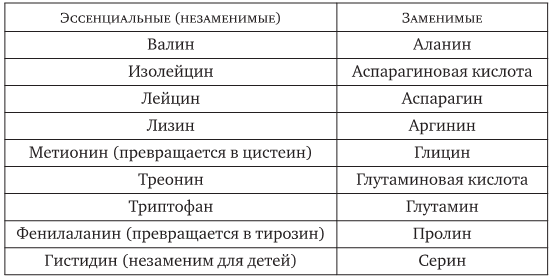 Аминокислоты
2009 г.; 37: 187–98. [PubMed] [Google Scholar]
Аминокислоты
2009 г.; 37: 187–98. [PubMed] [Google Scholar]
24. Liu XD, Wu X, Yin YL, Liu YQ, Geng MM, Yang HS, Blachier F, Wu GY. Влияние пищевых добавок L-аргинина или N-карбамилглутамата на поздних сроках беременности свиноматок на экспрессию miR-15b/16, miR-221/222, VEGFA и eNOS в пупочной вене. Аминокислоты 2012 г.; 42: 2111–9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Wang WW, Wu ZL, Lin G, Hu SD, Wang B, Dai ZL, Wu G. Глицин стимулирует синтез белка и ингибирует окислительный стресс в эпителиальных клетках тонкого кишечника свиней. Джей Нутр 2014; 144: 1540–158. [PubMed] [Google Scholar]
26. Rhoads JM, Niu XM, Surendran S, Liu YY, Wu G. Аргинин стимулирует миграцию эпителиальных клеток кишечника, трансформированных cdx2, посредством механизма, требующего передачи сигналов как оксидом азота, так и p70s6k. Джей Нутр 2008 г.; 138: 1652–167. [PubMed] [Академия Google]
27. Wang XQ, Frank JW, Xu J, Dunlap KA, Satterfield MC, Burghardt RC, Romero JJ, Hansen TR, Wu G, Bazer FW. Функциональная роль аргинина в периимплантационный период беременности. II. Последствия потери функции мРНК синтазы оксида азота NOS3 в трофэктодерме концептуса овцы. Биол Репрод
2014; 91: 1–10. [PubMed] [Google Scholar]
Функциональная роль аргинина в периимплантационный период беременности. II. Последствия потери функции мРНК синтазы оксида азота NOS3 в трофэктодерме концептуса овцы. Биол Репрод
2014; 91: 1–10. [PubMed] [Google Scholar]
28. Yao K, Yin YL, Chu WY, Liu ZQ, Deng D, Li TJ, Huang RL, Zhang JS, Tan BE, Wang W, Wu G. Добавка аргинина в пищу увеличивает сигнальную активность mTOR в скелетных мышцах новорожденных свиней. Джей Нутр 2008 г.; 138: 867–872. [PubMed] [Академия Google]
29. Ван Х., Чжан С., У Г., Сунь Ю.Л., Ван Б., Хе Б.Б., Дай З.Л., Ву З.Л. Глутамин усиливает экспрессию белка плотных контактов и модулирует передачу сигналов CRF в тощей кишке поросят-отъемышей. Джей Нутр 2015 г.; 145: 25–31. [PubMed] [Google Scholar]
30. Zhang J, Yin YL, Shu XG, Li TJ, Li FN, Tan BE, Wu ZL, Wu G.
Пероральное введение глутамата натрия увеличивает экспрессию глутаматных рецепторов и транспортеров в желудочно-кишечном тракте молодых поросят. Аминокислоты
2013; 45: 1169–1177. [PubMed] [Академия Google]
[PubMed] [Академия Google]
31. Сан Габриэль А., Унеяма Х. Чувствительность аминокислот в желудочно-кишечном тракте. Аминокислоты 2013; 45: 451–61. [PubMed] [Google Scholar]
32. Dai ZL, Wu G, Zhu WY. Метаболизм аминокислот в кишечных бактериях: связь между экологией кишечника и здоровьем хозяина. Фронт БиоСки 2011 г.; 16: 1768–1786. [PubMed] [Google Scholar]
33. Wang WW, Dai ZL, Wu ZL, Lin G, Jia SC, Hu SD, Dahanayaka S, Wu G. Глицин является питательно незаменимой аминокислотой для максимального роста молодняка молочных свиней. Аминокислоты 2014; 46: 2037–45. [PubMed] [Академия Google]
34. Дай ZL, Li XL, Xi PB, Zhang J, Wu G, Zhu WY. L-глютамин регулирует использование аминокислот кишечными бактериями. Аминокислоты 2013; 45: 501–12. [PubMed] [Google Scholar]
35. Dai ZL, Li XL, Xi PB, Zhang J, Wu G, Zhu WY. Регуляторная роль L-аргинина в утилизации аминокислот тонкокишечными бактериями свиней. Аминокислоты 2012 г.; 43: 233–44. [PubMed] [Google Scholar]
36. Fu WJ, Haynes TE, Kohli R, Hu J, Shi W, Spencer TE, Carroll RJ, Meininger CJ, Wu G.
Пищевые добавки с L-аргинином уменьшают жировую массу у диабетических жирных крыс Zucker. Джей Нутр
2005 г.; 135: 714–21. [PubMed] [Академия Google]
Fu WJ, Haynes TE, Kohli R, Hu J, Shi W, Spencer TE, Carroll RJ, Meininger CJ, Wu G.
Пищевые добавки с L-аргинином уменьшают жировую массу у диабетических жирных крыс Zucker. Джей Нутр
2005 г.; 135: 714–21. [PubMed] [Академия Google]
37. McKnight JR, Satterfield MC, Jobgen WS, Smith SB, Spencer TE, Meininger CJ, McNeal CJ, Wu G. Благотворное влияние L-аргинина на снижение ожирения: потенциальные механизмы и важные последствия для здоровья человека. Аминокислоты 2010 г.; 39: 349–57. [PubMed] [Google Scholar]
38. Jobgen WS, Fried SK, Fu WJ, Meininger CJ, Wu G. Регуляторная роль пути аргинин-оксид азота в метаболизме энергетических субстратов. Дж Нутр Биохим 2006 г.; 17: 571–88. [PubMed] [Академия Google]
39. Ву З.Л., Саттерфилд М.С., Базер Ф.В., Ву Г. Регуляция развития бурой жировой ткани и уменьшение количества белого жира с помощью L-аргинина. Curr Opin Clin Nutr Metab Care 2012 г.; 15: 529–38. [PubMed] [Google Scholar]
40. Tan B, Yin YL, Liu ZQ, Li XG, Xu HJ, Kong XF, Huang RL, Tang WJ, Shinzato I, Smith SB, Wu G. Пищевые добавки L-аргинина увеличивают мышечную массу и уменьшают жировую массу у свиней на доращивании и откорме. Аминокислоты
2009 г.; 37: 169–75. [PubMed] [Google Scholar]
Пищевые добавки L-аргинина увеличивают мышечную массу и уменьшают жировую массу у свиней на доращивании и откорме. Аминокислоты
2009 г.; 37: 169–75. [PubMed] [Google Scholar]
41. Haynes TE, Li P, Li XL, Shimotori K, Sato H, Flynn NE, Wang JJ, Knabe DA, Wu G. L-глютамин или L-аланил-L-глутамин предотвращают индуцированную оксидантами или эндотоксинами гибель неонатальных энтероцитов. Аминокислоты 2009 г.; 37: 131–42. [PubMed] [Google Scholar]
42. Rezaei R, Knabe DA, Tekwe CD, Dahanayaka S, Ficken MD, Fielder SE, Eide SJ, Lovering SL, Wu G. Пищевые добавки с глутаматом натрия безопасны и улучшают показатели роста поросят после отъема. Аминокислоты 2013; 44: 911–23. [PubMed] [Google Scholar]
43. Карнер С.М., Эсен Э., Окунаде А.Л., Паттерсон Б.В., Лонг Ф. Повышенный катаболизм глутамина опосредует анаболизм костей в ответ на передачу сигналов WNT. Джей Клин Инвест 2015 г.; 125: 551–62. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Ли П., Инь Ю.Л. , Ли Д.Ф., Ким С.В., Ву Г.
Аминокислоты и иммунная функция. Бр Дж Нутр
2007 г.; 98: 237–52. [PubMed] [Google Scholar]
, Ли Д.Ф., Ким С.В., Ву Г.
Аминокислоты и иммунная функция. Бр Дж Нутр
2007 г.; 98: 237–52. [PubMed] [Google Scholar]
45. Wu G, Flynn NE, Flynn SP, Jolly CA, Davis PK. Дефицит пищевого белка или аргинина ухудшает конститутивный и индуцируемый синтез оксида азота у молодых крыс. Джей Нутр 1999 г.; 129: 1347–1354. [PubMed] [Google Scholar]
46. De Jonge WJ, Kwikkers KL, Te Velde AA, Van Deventer SJH, Nolte MA, Mebius RE, Ruijter JM, Lamers MC, Lamers WH. Дефицит аргинина влияет на раннее созревание В-клеток и развитие лимфоидных органов у трансгенных мышей. Джей Клин Инвест 2002 г.; 110: 1539–48. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Ren WK, Liu SP, Chen S, Zhang FM, Li NZ, Yin J, Peng YY, Wu L, Liu G, Yin YL, Wu G. Пищевая добавка L-глютамина увеличивает бремя Pasteurella multocida и экспрессию основных факторов вирулентности у мышей. Аминокислоты 2013; 45: 947–55. [PubMed] [Google Scholar]
48. Ren WK, Zou LX, Ruan Z, Li NZ, Wang Y, Peng Y, Liu G, Yin YL, Huang RL, Hou YQ, Wu G. Пищевая добавка L-пролина оказывает иммуностимулирующее действие на инактивированные 9Мыши, иммунизированные вакциной Pasteurella multocida 0055. Аминокислоты
2013; 45: 555–61. [PubMed] [Google Scholar]
Пищевая добавка L-пролина оказывает иммуностимулирующее действие на инактивированные 9Мыши, иммунизированные вакциной Pasteurella multocida 0055. Аминокислоты
2013; 45: 555–61. [PubMed] [Google Scholar]
49. Bazer FW, Wu G, Johnson GA, Wang XQ. Факторы окружающей среды, влияющие на беременность: эндокринные разрушители, питательные вещества и метаболические пути. Мол Селл Эндокринол 2014; 398: 53–68. [PubMed] [Google Scholar]
50. Häberle J, Görg B, Rutsch F, Schmidt E, Toutain A, Benoist JF, Gelot A, Suc AL, Höhne W, Schliess F, Häussinger D, Koch HG. Врожденный дефицит глутамина с мутациями глутаминсинтетазы. N Engl J Med 2005 г.; 353: 1926–33. [PubMed] [Google Scholar]
51. Ву Г., Базер Ф.В., Дэвис Т.А., Ким С.В., Ли П., Роадс Дж.М., Саттерфилд М.С., Смит С.Б., Спенсер Т.Е., Инь Ю.Л. Метаболизм аргинина и питание при росте, здоровье и болезни. Аминокислоты 2009 г.; 37: 153–68. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Newsholme P, Abdulkader F, Rebelato E, Romanatto T, Pinheiro CH, Vitzel KF, Silva EP, Bazotte RB, Procopio J, Curi R, Gorjao R, Pithon -Кури ТС. Аминокислоты и диабет: последствия для эндокринной, метаболической и иммунной функции. Фронт БиоСки
2011 г.; 16: 315–39. [PubMed] [Google Scholar]
Аминокислоты и диабет: последствия для эндокринной, метаболической и иммунной функции. Фронт БиоСки
2011 г.; 16: 315–39. [PubMed] [Google Scholar]
53. Li C, Liu C, Nissim I, Chen J, Chen P, Doliba N, Zhang T, Nissim I, Daikhin Y, Stokes D, Yudkoff M, Bennett MJ, Stanley CA, Матчинский Ф.М., Наджи А. Регуляция секреции глюкагона в нормальных и диабетических островках человека с помощью γ-гидроксибутирата и глицина. J Биол Хим 2013; 288: 3938–51. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Hou YQ, Wang L, Yi D, Wu G. N-ацетилцистеин и здоровье кишечника: в центре внимания механизмы его действия. Фронт БиоСки 2015 г.; 20: 872–8791. [PubMed] [Google Scholar]
55. Цуругизава Т., Унеяма Х., Тории К. Восприятие аминокислот мозгом. Диабет Ожирение Метаб 2014; 16: 41–8. [PubMed] [Google Scholar]
56. Billard JM. D-аминокислоты в нейротрансмиссии мозга и синаптической пластичности. Аминокислоты 2012 г.; 43: 1851–1860. [PubMed] [Google Scholar]
57. Руццо Э.К., Капо-Чичи Дж.М., Бен-Зеев Б., Читаят Д., Мао Х., Паппас А.Л., Хитоми Ю., Лу Ю.Ф., Яо Х., Хамдан Ф.Ф., Пелак К., Резник- Вольф Х., Бар-Джозеф И., Оз-Леви Д., Лев Д., Лерман-Саги Т., Лешински-Сильвер Э., Аникстер Ю., Бен-Ашер Э., Олендер Т., Колло Л., Декари Дж. К., Блазер С., Банвелл Б., Джоши RB, He XP, Patry L, Silver RJ, Dobrzeniecka S, Islam MS, Hasnat A, Samuels ME, Aryal DK, Rodriguiz RM, Jiang YH, Wetsel WC, McNamara JO, Rouleau GA, Silver DL, Lancet D, Pras E, Митчелл Г.А., Мишо Дж.Л., Гольдштейн Д.Б.
Дефицит аспарагинсинтетазы вызывает врожденную микроцефалию и прогрессирующую форму энцефалопатии. Нейрон
2013; 80: 429–41. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Руццо Э.К., Капо-Чичи Дж.М., Бен-Зеев Б., Читаят Д., Мао Х., Паппас А.Л., Хитоми Ю., Лу Ю.Ф., Яо Х., Хамдан Ф.Ф., Пелак К., Резник- Вольф Х., Бар-Джозеф И., Оз-Леви Д., Лев Д., Лерман-Саги Т., Лешински-Сильвер Э., Аникстер Ю., Бен-Ашер Э., Олендер Т., Колло Л., Декари Дж. К., Блазер С., Банвелл Б., Джоши RB, He XP, Patry L, Silver RJ, Dobrzeniecka S, Islam MS, Hasnat A, Samuels ME, Aryal DK, Rodriguiz RM, Jiang YH, Wetsel WC, McNamara JO, Rouleau GA, Silver DL, Lancet D, Pras E, Митчелл Г.А., Мишо Дж.Л., Гольдштейн Д.Б.
Дефицит аспарагинсинтетазы вызывает врожденную микроцефалию и прогрессирующую форму энцефалопатии. Нейрон
2013; 80: 429–41. [Бесплатная статья PMC] [PubMed] [Google Scholar]
58. Curthoys NP, Watford M. Регуляция активности глутаминазы и метаболизма глутамина. Анну Рев Нутр 1995 год; 15: 133–59. [PubMed] [Google Scholar]
59. Taylor L, Curthoys NP. Метаболизм глютамина: роль в кислотно-щелочном балансе. Биохим Мол Биол Эдук 2004 г.; 32: 291–304. [PubMed] [Google Scholar]
60. Штурм Р.А.
Молекулярная генетика разнообразия пигментации человека. Хум Мол Жене
2009 г.; 18: Р9–17. [PubMed] [Академия Google]
Штурм Р.А.
Молекулярная генетика разнообразия пигментации человека. Хум Мол Жене
2009 г.; 18: Р9–17. [PubMed] [Академия Google]
61. Ланг Ф., Буш Г.Л., Риттер М., Фёлькль Х., Вальдеггер С., Гулбинс Э., Хауссингер Д. Функциональное значение механизмов регуляции клеточного объема. Физиол Преподобный 1998 год; 78: 247–306. [PubMed] [Google Scholar]
62. Marliss EB, Aoki TT, Pozefsky T, Most AS, Cahill GF., Jr. Мышечный и внутренностный метаболизм глутамина и глутамата у постабсорбтивного и голодающего человека. Джей Клин Инвест 1971 год; 50: 814–7. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Wu G, Meininger CJ, Knabe DA, Bazer FW, Rhoads JM. Аргининовое питание в развитии, здоровье и болезни. Curr Opin Clin Nutr Metab Care 2000 г.; 3: 59–66. [PubMed] [Google Scholar]
64. Guedes RLM, Prosdocimi F, Fernandes GR, Moura LK, Ribeiro HAL, Ortega JM.
Биосинтез аминокислот и пути ассимиляции азота: большая геномная делеция в ходе эволюции эукариот. Геномика BMC
2011 г. ; 12: С2–С2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
; 12: С2–С2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Holt LE, Jr, Albanese AA. Наблюдения за дефицитом аминокислот у человека. Trans Assoc Am Врачи 1944 год; 58: 143–56. [Google Scholar]
66. Мейстер А. Биохимия аминокислот, Нью-Йорк: Academic Press, 19.65. [Google Scholar]
67. Танимура Дж. Исследования аргинина в сперме человека. Часть II. Влияние лекарств с L-аргинином-HCl на мужское бесплодие. Медицинская школа Булла в Осаке 1967 год; 13: 84–9. [PubMed] [Google Scholar]
68. Мелендес-Эвиа Э., Де Пас-Луго П., Корниш-Боуден А., Карденас М.Л. Слабое звено метаболизма: метаболические возможности биосинтеза глицина не удовлетворяют потребность в синтезе коллагена. Джей Биоски 2009 г.; 34: 853–72. [PubMed] [Google Scholar]
69. Севастиаду С., Маламици-Пухнер А., Косталос С., Скуролиаку М., Бриана Д. Д., Анцаклис А., Рома-Яннику Э.
Влияние перорального приема глютамина на проницаемость кишечника и частоту развития некротизирующего энтероколита/септицемии у недоношенных новорожденных. J Matern Fetal Neonatal Med
2011 г.; 24: 1294–300. [PubMed] [Google Scholar]
J Matern Fetal Neonatal Med
2011 г.; 24: 1294–300. [PubMed] [Google Scholar]
70. Rogers QR, Chen DM, Harper AE. Важность незаменимых аминокислот для максимального роста крыс. Proc Soc Exp Biol Med 1970 г.; 134: 517–22. [PubMed] [Google Scholar]
71. Breuer LH, Jr, Pond WG, Warner RG, Loosli JK. Роль незаменимых аминокислот в питании крысы. Джей Нутр 1964 год; 82: 499–506. [PubMed] [Google Scholar]
72. Вомак М. Исследования кормления аминокислотами: влияние различных заменимых источников азота и добавленной воды. Proc Soc Exp Biol Med 1969; 131: 977–9. [PubMed] [Google Scholar]
73. Stucki WP, Harper AE. Значение незаменимых аминокислот для нормального роста цыплят. Джей Нутр 1961 год; 74: 377–83. [Google Scholar]
74. Маруяма К., Сунде М.Л., Харпер А.Е. Является ли L-глутаминовая кислота незаменимой аминокислотой для молодых цыплят? Науки о птицеводстве 1976 год; 55: 45–60. [PubMed] [Google Scholar]
75. Wu G, Bazer FW, Johnson GA, Knabe DA, Burghardt RC, Spencer TE, Li XL, Wang JJ. Важная роль L-глютамина в кормлении и производстве свиней. Джей Аним Наука
2011 г.; 89: 2017–30. [PubMed] [Google Scholar]
Важная роль L-глютамина в кормлении и производстве свиней. Джей Аним Наука
2011 г.; 89: 2017–30. [PubMed] [Google Scholar]
76. Lei J, Feng DY, Zhang YL, Zhao FQ, Wu ZL, San Gabriel A, Fujishima Y, Uneyama H, Wu G. Питательная и регуляторная роль аминокислот с разветвленной цепью в лактации. Фронт БиоСки 2012 г.; 17: 2725–39. [PubMed] [Google Scholar]
77. Deng D, Yin YL, Chu WY, Yao K, Li TJ, Huang RL, Liu ZQ, Zhang JS, Wu G. Нарушение активации инициации трансляции и снижение синтеза белка у поросят-отъемышей, получавших рацион с низким содержанием белка. Дж Нутр Биохим 2009 г.; 20: 544–52. [PubMed] [Академия Google]
78. Дэвис Т.А., Фиоротто М.Л. Регуляция роста мышц у новорожденных. Curr Opin Nutr Metab Care 2009 г.; 12: 78–85. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Wu G. Пищевые потребности животных в синтезируемых аминокислотах: изменение парадигмы белкового питания. J Anim Sci Biotechnol 2014; 5: 34–34. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Li XL, Rezaei R, Li P, Wu G.
Состав аминокислот в кормовых ингредиентах для рационов животных. Аминокислоты
2011 г.; 40: 1159–68. [PubMed] [Google Scholar]
Li XL, Rezaei R, Li P, Wu G.
Состав аминокислот в кормовых ингредиентах для рационов животных. Аминокислоты
2011 г.; 40: 1159–68. [PubMed] [Google Scholar]
81. Dai ZL, Wu ZL, Jia SC, Wu G. Анализ аминокислотного состава белков животных тканей и пищевых продуктов в виде предколоночных производных o -фталдиальдегида методом ВЭЖХ с флуоресцентным детектированием. J Хроматогр B 2014; 964: 116–27. [PubMed] [Google Scholar]
82. Hou Y, Wang L, Zhang W, Yang ZG, Ding BY, Zhu HL, Liu YL, Qiu YS, Yin YL, Wu G. Защитное действие N-ацетилцистеина на функции кишечника поросят, зараженных липополисахаридом. Аминокислоты 2012 г.; 43: 1233–42. [PubMed] [Академия Google]
Действительно ли незаменимые аминокислоты не нужны?
Vetagro Journal Club, #5
Аминокислоты (АК) традиционно подразделяются на незаменимые и заменимые в зависимости от способности организма синтезировать их ex novo. Тем не менее, это определение частично неверно, поскольку некоторые так называемые «незаменимые» АК (ЭАК) могут быть синтезированы из предшественников, которые химически и структурно схожи. Например, метионин может быть синтезирован переаминированием его кетокислоты, а также деметилированием гомоцистеина, и в этом отношении единственными действительно незаменимыми АК могут быть треонин и лизин.
Например, метионин может быть синтезирован переаминированием его кетокислоты, а также деметилированием гомоцистеина, и в этом отношении единственными действительно незаменимыми АК могут быть треонин и лизин.
С другой стороны, в соответствии со строго метаболическим определением, настоящая заменимая аминокислота (NEAA) должна быть синтезирована de novo из неаминокислотного источника азота и углерода: в данном случае только глутаминовой кислоты и серина будет классифицироваться как действительно NEAA.
Существуют некоторые АК, которые могут быть синтезированы более сложными путями , чем простое трансаминирование соответствующей кетокислоты, и они называются условно незаменимыми, поскольку существуют постоянные ограничения в отношении степени и времени, когда эти АК могут быть синтезированы.
В основном считается, что потребность в АК диктуется накоплением и развитием белка, но на самом деле это составляет только 10% от общей потребности в АК. На самом деле AA участвуют в большинстве наиболее важных физиологических реакций , таких как регуляция экспрессии генов среди прочих. В частности, глутамин, глутамат, аргинин, глицин и пролин, которые классифицируются как NEAA, вовлечены в различные метаболические функции, такие как путь mTOR для синтеза белка в скелетных мышцах; синтез оксида азота, участвующий в противовоспалительном ответе, передачу сигналов клетками через цАМФ или цГМФ-зависимые киназы и многие другие. NEAA также может быть предшественник других биоактивных молекул , таких как карнозин, креатин, глутатион, полиамины и нейротрансмиттеры , которые являются важными регуляторами роста и дифференцировки клеток и развития тканей.
В частности, глутамин, глутамат, аргинин, глицин и пролин, которые классифицируются как NEAA, вовлечены в различные метаболические функции, такие как путь mTOR для синтеза белка в скелетных мышцах; синтез оксида азота, участвующий в противовоспалительном ответе, передачу сигналов клетками через цАМФ или цГМФ-зависимые киназы и многие другие. NEAA также может быть предшественник других биоактивных молекул , таких как карнозин, креатин, глутатион, полиамины и нейротрансмиттеры , которые являются важными регуляторами роста и дифференцировки клеток и развития тканей.
На сегодняшний день пищевые потребности АК у разных видов установлены только для EAA, но это классическое различение по своей природе имеет слабые стороны, поскольку в конкретных физиологических контекстах потребности организма в определенных АК могут быть увеличены сверх его возможностей синтезировать их .
Поскольку последние данные подтверждают необходимость установления требований NEAA, мы должны тщательно пересмотреть наш подход к белковому/аминокислотному питанию как для скота, так и для людей, поскольку мы можем недооценивать реальные потребности, особенно для тех категорий, которые в большей степени подвержены специфическим потребности в питании, такие как матери во время беременности и кормления грудью, недоношенные дети, профессиональные спортсмены и пожилые люди.
