ИВЭПР 12/3,5 RS-R3
ИВЭПР 12/3,5 RS-R3Арт.: RBZ-216602
ИСБ RUBEZH R3 Системы противопожарной защиты Питание
Предназначен для стабильного бесперебойного электропитания устройств охранно-пожарной сигнализации с напряжением питания 12 В постоянного тока.
8 294 ₽* /шт.
* Цена базового исполнения
Где купить?
Калькулятор
Назначение:
Источник обеспечивает стабильное круглосуточное электропитание оборудования систем охранной и пожарной сигнализации номинальным напряжением 12 В постоянным током до 3,5 А. Для увеличения времени резервирования к источнику может подключаться необходимое количество внешних боксов резервного питания БР 12 производства «Рубеж».
Источник обеспечивает формирование и передачу в приемно-контрольные приборы Рубеж по адресной линии связи по протоколу RS-R3 следующие сообщения:
События:
- Разряд АКБ1
- Разряд АКБ1 устранён
- Отсутствие АКБ1
- АКБ1 подключен
- Нет сетевого напряжения
- Разряд АКБ2
- Разряд АКБ2 устранён
- Отсутствие АКБ2
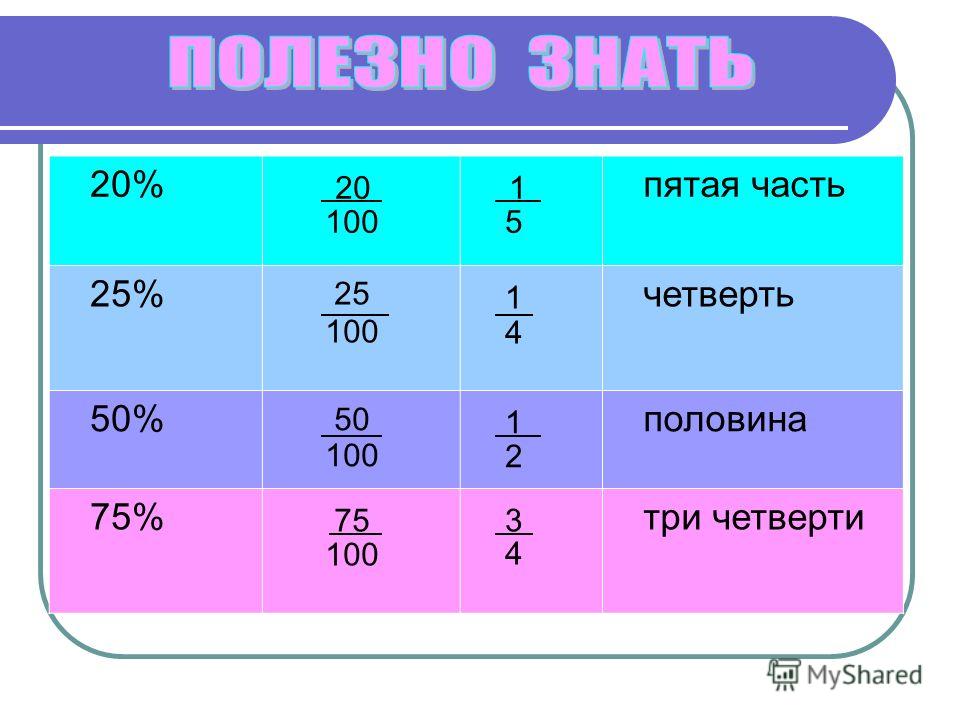
- АКБ2 подключен
- Есть сетевое напряжение
Неисправности:
- Отсутствие напряжения на выходе, напряжение < 9В (выход 1)
- Отсутствие напряжения на выходе, напряжение < 9В (выход 2)
- Глубокий разряд АКБ1
- Глубокий разряд АКБ2
Источник также передает в ППКП текущие значения выходного напряжения по каждому выходу, а также напряжения каждой из двух АКБ.
Преимущества конструкции:
- оптимальный заряд АКБ за счёт схемы постоянного тестирования;
- защиту аккумуляторных батарей от перезаряда и глубокого разряда.
- электронная защита от переполюсовки при подключении АКБ и замыкания клемм присоединения АКБ. При работе от сети переполюсовка или замыкание клемм АКБ не влияет на выходное напряжение;
- электронная защита источника от короткого замыкания и перегрузок по току, как при работе от сети, так и при работе от АКБ. После устранения режима короткого замыкания или перегрузки источник автоматически восстанавливает выходное напряжение.
Схемное решение:
- возможность подключения адресной линии связи для передачи информации о состоянии источника питания;
- автоматический заряд АКБ при наличии сети переменного тока;
- автоматический переход на работу от АКБ при снижении сетевого напряжения ниже минимального рабочего значения и возврат на питание от сети при восстановлении сетевого напряжения;
- низкий уровень высокочастотных помех за счет оптимальной топологии печатной платы;
- выходное напряжение гальванически развязано от корпуса.

В источнике предусмотрен сигнал «Авария» для передачи информации о наличии неисправности во внешние цепи в виде электромагнитного реле. Выход сигнала «Авария» изолирован от цепей источника и корпуса. При одновременном наличии сетевого напряжения, хотя бы одной исправной и заряженной АКБ, и выходного напряжения – реле замкнуто. Иначе – разомкнуто.
Напряжение питания от сети переменного тока, В
140 – 265
Мощность, потребляемая от сети переменного тока, Вт, не более
80
Собственное потребление источника от АКБ в резервном режиме, мА, не более
30
Выходное напряжение источника при работе от сети, В
13,3 — 13,8
Выходное напряжение источника при работе от АКБ, В
9,4 — 13,5
Количество устанавливаемых АКБ
2
Емкость АКБ, А·ч
от 7 до 17
Максимальный ток нагрузки, обеспечиваемый источником, А, не более
3,5
Масса (без АКБ), кг, не более
см. таблицу исполнений
Размеры
см. таблицу исполнений
таблицу исполненийГарантийный срок эксплуатации, мес, не более
24
Средний срок службы, лет
10
Диапазон рабочих температур, °С
от -10 до +50
| Параметр | Значение |
|---|---|
| Напряжение питания от сети переменного тока, В | 140 – 265 |
| Мощность, потребляемая от сети переменного тока, Вт, не более | 80 |
| Собственное потребление источника от АКБ в резервном режиме, мА, не более | 30 |
| Выходное напряжение источника при работе от сети, В | 13,3 — 13,8 |
| Выходное напряжение источника при работе от АКБ, В | 9,4 — 13,5 |
| Количество устанавливаемых АКБ | 2 |
| Емкость АКБ, А·ч | от 7 до 17 |
| Максимальный ток нагрузки, обеспечиваемый источником, А, не более | 3,5 |
| Масса (без АКБ), кг, не более | см. таблицу исполнений таблицу исполнений |
| Размеры | см. таблицу исполнений |
| Гарантийный срок эксплуатации, мес, не более | 24 |
| Средний срок службы, лет | 10 |
| Диапазон рабочих температур, °С | от -10 до +50 |
Источники вторичного электропитания резервированные ИВЭПР 12, в зависимости от исполнения, могут иметь следующие обозначения:
1 – Наименование изделия: ИВЭПР – источник вторичного электропитания резервированный.
2 – Выходное напряжение: 12 – 12 В.
3 – Номинальный выходной ток: 2 – 2 А, 3,5 – 3,5 А, 5 – 5 А.
4 – Работа в адресной системе: RS-R3 – для подключения в АЛС приборов протокола R3.
5 – Количество батарей: 2 – 2 АКБ.
6 – Максимальная емкость каждой батареи: 7 – 7 А*ч, 12 – 12 А*ч, 17 – 17 А*ч, 40 – 40 А*ч.
7 – Клеммы для подключения боксов резервного питания БР: БР – с клеммами.
Источники вторичного электропитания резервированные ИВЭПР 12/3,5 RS-R3 выпускаются в следующих исполнениях:
|
|
Количество и максимальная ёмкость АКБ |
Размеры корпуса, мм |
Масса без АКБ, кг (не более) |
Возможность подключения боксов БР12 |
|
ИВЭПР 12/3,5 RS-R3 2х7 БР |
2×7 А*ч | 324×86×184 | 2,1 |
Есть |
|
ИВЭПР 12/3,5 RS-R32х12 БР |
2×12 А*ч |
324×111×184 |
2,3 | Есть |
|
ИВЭПР 12/3,5 RS-R3 2х17 БР |
2×17 А*ч | 254×376×86 | 2,6 | Есть |
- Сертификат соответствия ТР ЕАЭС 043/2017 (PDF, 763.
 1 КБ)
1 КБ) - Декларация (PDF, 822.18 КБ)
- Сертификат соответствия постановлению правительства РФ № 969 от 26.09.2016 (PDF, 4.28 МБ)
- Паспорт (PDF, 2.29 МБ)
- Инструкция по монтажу (PDF, 1.4 МБ)
- Инструкция по подключению (PDF, 1.37 МБ)
- Схема подключения (DWG, 62.84 КБ)
Огнетушитель ОУ-3 (5 литров) углекислотный цена в Санкт-Петерубрге: 1694 руб.
Перейти к основному содержанию Skip to navigation
Вход для клиентов
07 марта 2023 С Праздником 8 Марта!
В наличии
Розничная
1 694 р.
Оптовая
1 590 р.
Описание
Огнетушитель ОУ-3 — переносной углекислотный огнетушитель с массой заряда огнетушащего вещества (ОТВ) 3 кг.
Баллон огнетушителя наполнен двуокисью углерода и находится под давлением.
Огнетушитель ОУ-3 необходимо перезаряжать после использования, либо по истечении срока 5 лет.
Огнетушитель углекислотный ОУ-3 предназначен для защиты промышленных и гражданских объектов в качестве первичного средства тушения возгораний.
Подробнее
Табы
5 причин работать с намиISO 9001-2011
Наша компания сертифицирована по системе менеджмента качества ISO 9001-2011. № РОСС RU.OC04.CMK.00278 JN 09.09.2015г.Качество продукции
Мы гарантируем отсутствие брака.Лучшая цена
Мы умеем продавать дешевле! Поставки напрямую от производителя и скидки для постоянных покупателейАссортимент и наличие
Мы не работаем «под заказ» — вся продукция в наличии на нашем оптовом складе в Санкт-Петербурге.Оперативность
Если вам нужно «вчера», а сегодня уже приходит инспектор госпожнадзора с проверкой, звоните нам!Профессиональная команда
Каждый сотрудник проходит аттестацию в области пожарной безопасности. Характеристики
ХарактеристикиТехнические характеристики Огнетушителя ОУ-3 BCE (5 литров) d=114 мм
| Производство | Ярпожинвест |
|---|---|
| Масса заряда ОТВ | 3.00кг |
| Температура эксплуатации и хранения | от -40°С до 50°С |
| Габаритные размеры | 600 х 114 мм |
| Вес | 10,5 кг |
| Время выхода ОТВ | 8 секунд |
| Длина выброса ОТВ | 3 м |
| Возможность перезарядки | есть |
| Срок службы до перезарядки | 5 лет |
| Гарантийный срок эксплуатации заряженного огнетушителя | 1 год |
| Ранг модельного очага | 34В |
| Применение | Огнетушитель для дома Огнетушители для офиса Огнетушитель для техосмотра Огнетушитель для электроустановок |
Применение ОУ-3
Огнетушитель углекислотный ОУ-3 ВСЕ применяется для тушения в соответствии с классами пожара:
- В — пожары горючих жидкостей или плавящихся твердых веществ и материалов;
- С — пожары газов;
- Е — пожары горючих веществ и материалов электроустановок, находящихся под напряжением.

Огнетушитель углекислотный ОУ-3 НЕ ПРЕДНАЗНАЧЕН для тушения возгораний, соответствующих классу пожара А — пожары твердых горючих веществ и материалов.
Возможны незначительные отклонения веса и габаритных размеров от указанных по причине использования производителями различных баллонов.
Преимущество углекислотных огнетушителей в том, что огнетушащий состав не причиняет вреда объекту тушения и обладает хорошими диэлектрическими свойствами. Главным достоинством углекислотных огнетушителей является их особенность не портить при тушении электронное оборудование, приборы и механизмы, а также и само охраняемое помещение.
Углекислотными огнетушителями рекомендуется оснащать офисы и другие помещения с наличием оргтехники, транспортные средства, культурно-исторические объекты и архивы.
Огнетушители ОУ не предназначены для тушения возгораний дерева и веществ, горение которых может происходить без доступа воздуха. При работе с углекислотным огнетушителем нужно соблюдать осторожность, так как при неаккуратном с ним обращении можно получить низкотемпературный ожог.
Использование переносного углекислотного огнетушителя ОУ-3:
- Поднести переносной углекислотный огнетушитель ОУ-3 к очагу пожара, с учетом безопасного от теплового воздействия расстояния.
- Выдернуть чеку, направить раструб на очаг загорания и нажать ручку клапана запорного устройства.
- В случаях загорании на открытом воздухе тушение производить только с наветренной стороны.
- Необходимо соблюдать осторожность при выпуске заряда двуокиси углерода из раструба, т.к. температура поверхности раструба с шлангом снижается до -60-70 °С.
- Кроме того, на поверхности раструба может концентрироваться электростатическое напряжение, способное пробить диэлектрическую перчатку.
- При тушении электрооборудования, находящегося под напряжением, не допускается подводить раструб или корпус огнетушителя ОУ-3 к открытым токоведущим частям с напряжением 10 кВ ближе чем на 2 метра.
УКАЗАНИЯ ПО ЭКСПЛУАТАЦИИ ОГНЕТУШИТЕЛЯ ОУ-3
- Огнетушитель ОУ-3 необходимо размещать в легкодоступных и заметных местах, где исключено попадание на него осадков, прямых солнечных лучей или нагрев огнетушителя выше 50 °С.

- Эксплуатация без чеки и пломбы завода изготовителя или организации, производившей перезарядку огнетушителя, не допускается.
- После применения огнетушитель ОУ-3 следует как можно быстрее отправить на перезарядку, заменив его однотипным резервным огнетушителем.
- Ежегодно необходимо проводить контрольное взвешивание огнетушителя при первоначальной установке и не реже одного раза в год. Допустимая величина утечки заряда двуокиси углерода -не более 50 г в год. При этом масса не должна выходить за переделы диапазона, казанного в пункте 2 табл.1.
- Перезарядка и техническое обслуживание огнетушителя должны проводиться не реже 1 раза в 5 лет специализированными организациями, имеющими лицензию на данный вид деятельности, с использованием специальной зарядной станции.
- Производить переосвидетельствование баллона через 10 лет.
Хранение
Для хранения необходимо использовать пожарные шкафы для огнетушителей, напольные подставки или осуществлять монтаж на вертикальную поверхность с помощью универсальных кронштейнов на высоте 1,5 м от уровня пола до верхней части корпуса.
Купить углекислотный огнетушитель ОУ-3 можно у нас в офисе или сделать предварительный заказ, заполнив форму заказа на сайте и оформить доставку.
9.2 Репликация ДНК – Концепции биологии – 1-е канадское издание
Перейти к содержанию
Глава 9: Введение в молекулярную биологию
К концу этого раздела вы сможете:
- Объяснять процесс репликации ДНК
- Объясните важность теломеразы для репликации ДНК
- Описать механизмы репарации ДНК
Когда клетка делится, важно, чтобы каждая дочерняя клетка получает идентичную копию ДНК . Это достигается за счет процесса репликации ДНК. Репликация ДНК происходит во время фазы синтеза или S-фазы клеточного цикла, прежде чем клетка вступает в митоз или мейоз.
Выяснение структуры двойной спирали дало представление о том, как копируется ДНК. Напомним, что адениновые нуклеотиды сочетаются с тиминовыми нуклеотидами, а цитозин с гуанином. Это означает, что две нити дополняют друг друга. Например, цепь ДНК с последовательностью нуклеотидов AGTCATGA будет иметь комплементарную цепь с последовательностью TCAGTACT (рис. 9)..8).
Напомним, что адениновые нуклеотиды сочетаются с тиминовыми нуклеотидами, а цитозин с гуанином. Это означает, что две нити дополняют друг друга. Например, цепь ДНК с последовательностью нуклеотидов AGTCATGA будет иметь комплементарную цепь с последовательностью TCAGTACT (рис. 9)..8).
Из-за комплементарности двух нитей наличие одной нити означает возможность воссоздания другой нити . Эта модель репликации предполагает, что две нити двойной спирали расходятся во время репликации, и каждая нить служит шаблоном, с которого копируется новая комплементарная нить (рис. 9)..9).
Рис. 9.9 Показана полуконсервативная модель репликации ДНК. Серым цветом обозначены исходные цепи ДНК, а синим — вновь синтезированная ДНК. Во время репликации ДНК каждая из двух цепей, составляющих двойную спираль, служит шаблоном, с которого копируются новые цепи. Новая цепь будет комплементарна родительской или «старой» цепи. Каждая новая двойная цепь состоит из одной родительской цепи и одной новой дочерней цепи. Это известно как полуконсервативная репликация. При образовании двух копий ДНК они имеют одинаковую последовательность нуклеотидных оснований и поровну делятся на две дочерние клетки.
Новая цепь будет комплементарна родительской или «старой» цепи. Каждая новая двойная цепь состоит из одной родительской цепи и одной новой дочерней цепи. Это известно как полуконсервативная репликация. При образовании двух копий ДНК они имеют одинаковую последовательность нуклеотидных оснований и поровну делятся на две дочерние клетки.
Поскольку эукариотические геномы очень сложны, репликация ДНК представляет собой очень сложный процесс, в котором участвуют несколько ферментов и других белков. Он протекает в три основных этапа: инициация, элонгация и терминация.
Напомним, что эукариотическая ДНК связана с белками, известными как гистоны, и образует структуры, называемые нуклеосомами. Во время инициации ДНК становится доступной для белков и ферментов, участвующих в процессе репликации. Как механизм репликации узнает, где начинается двойная спираль ДНК? Оказывается, существуют определенные последовательности нуклеотидов, называемые точками начала репликации, с которых начинается репликация. Определенные белки связываются с точкой начала репликации, в то время как фермент, называемый геликазой, раскручивает спираль ДНК. Когда ДНК раскрывается, образуются Y-образные структуры, называемые репликационными вилками (рис. 9)..10). В начале репликации формируются две вилки репликации, которые расширяются в обоих направлениях по мере репликации. На эукариотической хромосоме существует несколько источников репликации, так что репликация может происходить одновременно из нескольких мест в геноме.
Определенные белки связываются с точкой начала репликации, в то время как фермент, называемый геликазой, раскручивает спираль ДНК. Когда ДНК раскрывается, образуются Y-образные структуры, называемые репликационными вилками (рис. 9)..10). В начале репликации формируются две вилки репликации, которые расширяются в обоих направлениях по мере репликации. На эукариотической хромосоме существует несколько источников репликации, так что репликация может происходить одновременно из нескольких мест в геноме.
Во время элонгации фермент под названием ДНК-полимераза добавляет нуклеотиды ДНК к 3′-концу матрицы. Поскольку ДНК-полимераза может добавлять новые нуклеотиды только в конце остова, последовательность праймеров, которая обеспечивает эту отправную точку, добавляется с комплементарными нуклеотидами РНК. Позже этот праймер удаляют, а нуклеотиды заменяют нуклеотидами ДНК. Одна цепь, комплементарная родительской цепи ДНК, непрерывно синтезируется в направлении репликационной вилки, поэтому полимераза может добавлять нуклеотиды в этом направлении. Эта непрерывно синтезируемая цепь известна как ведущая цепь. Поскольку ДНК-полимераза может синтезировать ДНК только в направлении от 5′ к 3′, другая новая цепь собирается в короткие фрагменты, называемые фрагментами Окадзаки. Каждому из фрагментов Окадзаки для начала синтеза требуется праймер, состоящий из РНК. Цепь с фрагментами Окадзаки известна как отстающая нить. В ходе синтеза фермент удаляет РНК-праймер, который затем заменяется нуклеотидами ДНК, а промежутки между фрагментами заделываются ферментом, называемым ДНК-лигазой.
Эта непрерывно синтезируемая цепь известна как ведущая цепь. Поскольку ДНК-полимераза может синтезировать ДНК только в направлении от 5′ к 3′, другая новая цепь собирается в короткие фрагменты, называемые фрагментами Окадзаки. Каждому из фрагментов Окадзаки для начала синтеза требуется праймер, состоящий из РНК. Цепь с фрагментами Окадзаки известна как отстающая нить. В ходе синтеза фермент удаляет РНК-праймер, который затем заменяется нуклеотидами ДНК, а промежутки между фрагментами заделываются ферментом, называемым ДНК-лигазой.
Процесс репликации ДНК можно обобщить следующим образом:
- ДНК раскручивается в точке начала репликации.
- Новые основания добавляются к комплементарным родительским цепям. Одна новая прядь изготавливается непрерывно, а другая прядь изготавливается по частям.
- Удаляют праймеры, вместо праймеров помещают новые нуклеотиды ДНК и остов запечатывают с помощью ДНК-лигазы.
 Синтезируется РНК-праймер, который удлиняется ДНК-полимеразой. На ведущей цепи ДНК синтезируется непрерывно, тогда как на отстающей цепи ДНК синтезируется короткими отрезками. Фрагменты ДНК соединяют с помощью ДНК-лигазы (не показано).
Синтезируется РНК-праймер, который удлиняется ДНК-полимеразой. На ведущей цепи ДНК синтезируется непрерывно, тогда как на отстающей цепи ДНК синтезируется короткими отрезками. Фрагменты ДНК соединяют с помощью ДНК-лигазы (не показано).Вы изолируете клеточный штамм, в котором соединение фрагментов Оказаки нарушено, и подозреваете, что произошла мутация в ферменте, обнаруженном на репликационной вилке. Мутация какого фермента наиболее вероятна?
Репликация теломер
Поскольку эукариотические хромосомы линейны, репликация ДНК происходит в конце линии в эукариотических хромосомах. Как вы уже знаете, фермент ДНК-полимераза может присоединять нуклеотиды только в одном направлении. В ведущей цепи синтез продолжается до тех пор, пока не будет достигнут конец хромосомы; однако на отстающей цепи нет места для создания праймера для копируемого фрагмента ДНК на конце хромосомы. Это представляет проблему для клетки, потому что концы остаются непарными, и со временем эти концы становятся все короче, поскольку клетки продолжают делиться. Концы линейных хромосом известны как теломеры, которые имеют повторяющиеся последовательности, не кодирующие определенный ген. Как следствие, с каждым раундом репликации ДНК укорачиваются не гены, а теломеры. Например, у человека последовательность из шести пар оснований, TTAGGG, повторяется от 100 до 1000 раз. Открытие фермента теломеразы (рис. 9.11) помогли понять, как сохраняются концы хромосом. Теломераза прикрепляется к концу хромосомы, а на конце цепи ДНК добавляются основания, комплементарные матрице РНК. Как только матрица отстающей нити становится достаточно удлиненной, ДНК-полимераза теперь может добавлять нуклеотиды, комплементарные концам хромосом. Таким образом, концы хромосом реплицируются.
Концы линейных хромосом известны как теломеры, которые имеют повторяющиеся последовательности, не кодирующие определенный ген. Как следствие, с каждым раундом репликации ДНК укорачиваются не гены, а теломеры. Например, у человека последовательность из шести пар оснований, TTAGGG, повторяется от 100 до 1000 раз. Открытие фермента теломеразы (рис. 9.11) помогли понять, как сохраняются концы хромосом. Теломераза прикрепляется к концу хромосомы, а на конце цепи ДНК добавляются основания, комплементарные матрице РНК. Как только матрица отстающей нити становится достаточно удлиненной, ДНК-полимераза теперь может добавлять нуклеотиды, комплементарные концам хромосом. Таким образом, концы хромосом реплицируются.
Теломераза обычно активна в зародышевых клетках, взрослых стволовых клетках и некоторых раковых клетках. За открытие теломеразы и ее действия Элизабет Блэкберн (рис. 9.12) получила Нобелевскую премию по медицине и физиологии в 2009 году.
Теломераза не активна во взрослых соматических клетках. Взрослые соматические клетки, которые подвергаются клеточному делению, по-прежнему имеют укороченные теломеры. По сути, это означает, что укорочение теломер связано со старением. В 2010 году ученые обнаружили, что теломераза может обратить вспять некоторые возрастные состояния у мышей, и это может иметь потенциал в регенеративной медицине. 1 В этих исследованиях использовались мыши с дефицитом теломеразы; у этих мышей наблюдается атрофия тканей, истощение стволовых клеток, недостаточность системы органов и нарушение реакции на повреждение тканей. Реактивация теломеразы у этих мышей вызвала удлинение теломер, уменьшение повреждения ДНК, обратимую нейродегенерацию и улучшение функционирования яичек, селезенки и кишечника. Таким образом, реактивация теломер может иметь потенциал для лечения возрастных заболеваний у людей.
Репликация ДНК у прокариот
Напомним, что прокариотическая хромосома представляет собой кольцевую молекулу с менее обширной спиральной структурой, чем у эукариотических хромосом. Эукариотическая хромосома линейна и сильно закручена вокруг белков. Хотя в процессе репликации ДНК есть много общего, эти структурные различия обусловливают некоторые различия в процессе репликации ДНК у этих двух форм жизни.
Репликация ДНК чрезвычайно хорошо изучена у прокариот, в первую очередь из-за небольшого размера генома и большого количества доступных вариантов. Escherichia coli имеет 4,6 миллиона пар оснований в одной кольцевой хромосоме, и все они реплицируются примерно за 42 минуты, начиная с одного источника репликации и проходя по хромосоме в обоих направлениях. Это означает, что в секунду добавляется примерно 1000 нуклеотидов. Процесс идет намного быстрее, чем у эукариот. В таблице ниже приведены различия между прокариотическими и эукариотическими репликациями.
| Собственность | Прокариоты | Эукариоты |
|---|---|---|
| Происхождение репликации | Одноместный | Несколько |
| Скорость репликации | 1000 нуклеотидов/с | от 50 до 100 нуклеотидов/с |
| Хромосомная структура | циркулярный | линейный |
| Теломераза | Нет | Подарок |
Концепция в действии
Просмотрите руководство по репликации ДНК.
ДНК-полимераза может ошибаться при добавлении нуклеотидов. Он редактирует ДНК, проверяя каждую вновь добавленную базу. Неправильные базы удаляются и заменяются правильными, после чего полимеризация продолжается (рис. 9.13 a ). Большинство ошибок исправляются во время репликации, хотя, когда этого не происходит, используется механизм исправления несоответствия. Ферменты репарации несоответствия распознают неправильно включенное основание и вырезают его из ДНК, заменяя правильным основанием (рис. 9)..13 б ). При еще одном типе репарации, эксцизионной репарации нуклеотидов, двойная цепь ДНК раскручивается и разделяется, удаляются неправильные основания вместе с несколькими основаниями на 5′- и 3′-концах, которые заменяются путем копирования матрицы с помощью ДНК-полимеразы (рис. 9.13 c ). Эксцизионная репарация нуклеотидов особенно важна для коррекции димеров тимина, которые в первую очередь вызываются ультрафиолетовым светом. В димере тимина два нуклеотида тимина, соседствующие друг с другом на одной цепи, ковалентно связаны друг с другом, а не с комплементарными основаниями. Если димер не удалить и не восстановить, это приведет к мутации. Люди с дефектами в генах эксцизионной репарации нуклеотидов проявляют чрезвычайную чувствительность к солнечному свету и рано заболевают раком кожи.
Ферменты репарации несоответствия распознают неправильно включенное основание и вырезают его из ДНК, заменяя правильным основанием (рис. 9)..13 б ). При еще одном типе репарации, эксцизионной репарации нуклеотидов, двойная цепь ДНК раскручивается и разделяется, удаляются неправильные основания вместе с несколькими основаниями на 5′- и 3′-концах, которые заменяются путем копирования матрицы с помощью ДНК-полимеразы (рис. 9.13 c ). Эксцизионная репарация нуклеотидов особенно важна для коррекции димеров тимина, которые в первую очередь вызываются ультрафиолетовым светом. В димере тимина два нуклеотида тимина, соседствующие друг с другом на одной цепи, ковалентно связаны друг с другом, а не с комплементарными основаниями. Если димер не удалить и не восстановить, это приведет к мутации. Люди с дефектами в генах эксцизионной репарации нуклеотидов проявляют чрезвычайную чувствительность к солнечному свету и рано заболевают раком кожи.
 При восстановлении несоответствия (б) неправильно добавленная база обнаруживается после репликации. Белки восстановления несоответствия обнаруживают это основание и удаляют его из вновь синтезированной цепи под действием нуклеазы. Промежуток теперь заполнен правильно спаренной базой. Удаление нуклеотидов (с) восстанавливает димеры тимина. При воздействии УФ-излучения тимины, лежащие рядом друг с другом, могут образовывать димеры тимина. В нормальных клетках они вырезаются и замещаются.
При восстановлении несоответствия (б) неправильно добавленная база обнаруживается после репликации. Белки восстановления несоответствия обнаруживают это основание и удаляют его из вновь синтезированной цепи под действием нуклеазы. Промежуток теперь заполнен правильно спаренной базой. Удаление нуклеотидов (с) восстанавливает димеры тимина. При воздействии УФ-излучения тимины, лежащие рядом друг с другом, могут образовывать димеры тимина. В нормальных клетках они вырезаются и замещаются.Исправлено большинство ошибок; в противном случае они могут привести к мутации, определяемой как постоянное изменение в последовательности ДНК. Мутации в генах репарации могут привести к серьезным последствиям, таким как рак.
ДНКреплицируется полуконсервативным методом , в котором каждая из двух цепей родительской ДНК действует как матрица для синтеза новой ДНК. После репликации каждая ДНК имеет одну родительскую или «старую» цепь и одну дочернюю или «новую» цепь.
Репликация у эукариот начинается с нескольких источников репликации, в то время как репликация у прокариот начинается с одного источника репликации. ДНК вскрывается ферментами, в результате чего образуется репликационная вилка. Primase синтезирует РНК-праймер для инициации синтеза ДНК-полимеразой, которая может добавлять нуклеотиды только в одном направлении. Одна цепь синтезируется непрерывно в направлении репликационной вилки; это называется ведущей нитью. Другая цепь синтезируется в направлении, противоположном репликационной вилке, на коротких участках ДНК, известных как фрагменты Оказаки. Эта нить известна как отстающая нить. После завершения репликации РНК-праймеры заменяются нуклеотидами ДНК, а ДНК запечатывается с помощью ДНК-лигазы.
ДНК вскрывается ферментами, в результате чего образуется репликационная вилка. Primase синтезирует РНК-праймер для инициации синтеза ДНК-полимеразой, которая может добавлять нуклеотиды только в одном направлении. Одна цепь синтезируется непрерывно в направлении репликационной вилки; это называется ведущей нитью. Другая цепь синтезируется в направлении, противоположном репликационной вилке, на коротких участках ДНК, известных как фрагменты Оказаки. Эта нить известна как отстающая нить. После завершения репликации РНК-праймеры заменяются нуклеотидами ДНК, а ДНК запечатывается с помощью ДНК-лигазы.
Концы хромосом эукариот представляют собой проблему, так как полимераза не может удлинить их без праймера. Теломераза, фермент со встроенной матрицей РНК, удлиняет концы, копируя матрицу РНК и удлиняя один конец хромосомы. Затем ДНК-полимераза может удлинить ДНК с помощью праймера. Таким образом, концы хромосом защищены. В клетках есть механизмы для восстановления ДНК, когда она повреждается или при репликации возникают ошибки. Эти механизмы включают репарацию несоответствия для замены нуклеотидов, которые связаны с некомплементарным основанием, и эксцизионную репарацию нуклеотидов, которая удаляет поврежденные основания, такие как димеры тимина.
Эти механизмы включают репарацию несоответствия для замены нуклеотидов, которые связаны с некомплементарным основанием, и эксцизионную репарацию нуклеотидов, которая удаляет поврежденные основания, такие как димеры тимина.
Глоссарий
ДНК-лигаза: фермент, катализирующий соединение фрагментов ДНК вместе
ДНК-полимераза: фермент, синтезирующий новую цепь ДНК, комплементарную матричной цепи для открытия спирали ДНК во время репликации ДНК путем разрыва водородных связей
отстающая цепь: во время репликации 3′-5′-цепь, цепочка, которая реплицируется короткими фрагментами и удалена от репликационной вилки
ведущая цепь: цепь, которая непрерывно синтезируется в направлении от 5′ к 3′, которая синтезируется в направлении вилки репликации
репарация несоответствия: форма репарации ДНК, при которой распознаются, вырезаются и заменяются правильными нуклеотидами
мутация: постоянная вариация в последовательности нуклеотидов генома повреждения нуклеотидов, поврежденные нуклеотиды удаляются и заменяются новыми нуклеотидами с использованием комплементарной цепи, а цепь ДНК повторно запечатывается и позволяет воссоединиться с комплементарной
Фрагменты Оказаки: фрагменты ДНК, которые синтезируются короткими участками на отстающей цепи
праймер: короткий участок нуклеотидов РНК, который необходим для инициации репликации и позволяет ДНК-полимеразе связать и начать репликацию
репликация вилка: Y-образная структура, образующаяся во время инициации репликации
полуконсервативная репликация: метод, используемый для репликации ДНК, при котором двухцепочечная молекула разделяется, и каждая цепь действует как матрица для синтеза новой цепи , поэтому полученные молекулы ДНК состоят из одной новой нити нуклеотидов и одной старой нити нуклеотидов
теломераза: фермент, содержащий каталитическую часть и встроенную РНК-матрицу; он поддерживает теломеры на концах хромосом
теломеры: ДНК на концах линейных хромосом
Footnotes
1 Mariella Jaskelioff, et al. , «Реактивация теломеразы обращает вспять дегенерацию тканей у старых мышей с дефицитом теломеразы», Nature , 469 (2011):102–7.
, «Реактивация теломеразы обращает вспять дегенерацию тканей у старых мышей с дефицитом теломеразы», Nature , 469 (2011):102–7.
License
Concepts of Biology — 1st Canadian Edition Чарльза Молнара и Джейн Гейр распространяется по лицензии Creative Commons Attribution 4.0 International License, если не указано иное.
Поделиться этой книгой
Поделиться в Твиттере
Кибермост наук о жизни
Кибермост наук о жизниНуклеотиды и двойная спираль
ДНК, или дезоксирибонуклеиновая кислота, является наследственным материалом, присутствующим во всех клетках. ДНК дает инструкции по созданию, поддержанию и регулированию клеток и организмов и передается, когда клетки делятся и когда организмы размножаются. В этом блоке будет изучена молекулярная структура ДНК и ее упаковка внутри клеток. В 1953, используя данные, полученные Розалинд Франклин, Джеймс Уотсон и Фрэнсис Крик определили, что ДНК существует в форме, известной как двойная спираль. Спираль представляет собой извилистую структуру, похожую на штопор; ДНК известна как двойная спираль, потому что в каждой молекуле ДНК есть две переплетенные нити.
Спираль представляет собой извилистую структуру, похожую на штопор; ДНК известна как двойная спираль, потому что в каждой молекуле ДНК есть две переплетенные нити.
На изображении выше штопор показан слева с отмеченной спиральной областью. Изображение в центре показывает структуру ДНК. Обратите внимание, что есть две нити: одна показана синим, другая желтым. Другие примеры спирали включают пряжу, телефонный шнур или винтовую лестницу.
Каждая цепь двойной спирали состоит из повторяющихся звеньев, называемых нуклеотидами. Один нуклеотид состоит из трех функциональных групп: сахара, трифосфата и азотистого (азотсодержащего) основания, как показано ниже. Обратите внимание, что на рисунках, нарисованных в этом блоке, каждая непомеченная вершина структуры представляет собой атом углерода.
Сахар, обнаруженный в ДНК, представляет собой разновидность пятиуглеродного сахара, называемого рибозой. Структура рибозы изображена ниже. Каждый углерод рибозы пронумерован, как показано. Поскольку группа -ОН на 2′-углероде отсутствует в форме рибозы, обнаруженной в ДНК, сахар в ДНК называется 2′-дезоксирибозой.
Поскольку группа -ОН на 2′-углероде отсутствует в форме рибозы, обнаруженной в ДНК, сахар в ДНК называется 2′-дезоксирибозой.
Второй принципиальной особенностью нуклеотида является трифосфатная группа, присоединенная к 5′-углероду рибозной группы. В водной среде, как и внутри клетки, фосфатные группы заряжены отрицательно, как показано на рисунке выше.
Свободный невключенный нуклеотид обычно существует в форме трифосфата; то есть он содержит цепочку из трех фосфатов. Однако в ДНК она теряет две из этих фосфатных групп, так что только одна фосфатная группа включается в цепь ДНК. При включении нуклеотидов в ДНК соседние нуклеотиды соединяются фосфодиэфирной связью: между 5’-фосфатной группой одного нуклеотида и 3’-ОН-группой другого образуется ковалентная связь (см. ниже). Таким образом, каждая цепь ДНК имеет «остов» фосфат-сахар-фосфат-сахар-фосфат. Скелет имеет 5’-конец (со свободным фосфатом) и 3’-конец (со свободной ОН-группой). В приведенной ниже структуре каждый нуклеотид для ясности нарисован другим цветом.
Третьим принципиальным признаком нуклеотида является основание, которое присоединено к 1′-углероду рибозы. Хотя каждый нуклеотид в ДНК содержит идентичные сахарные и фосфатные группы, существует четыре разных основания и, следовательно, четыре разных нуклеотида, которые могут быть включены в ДНК. Четыре основания представляют собой аденин, цитозин, гуаин и тимин, и их структуры показаны ниже.
Когда эти основания входят в состав нуклеотидов, нуклеотиды называются 2’дезоксиаденозинтрифосфатом, 2’дезоксицитидинтрифосфатом, 2’дезоксигуанозинтрифосфатом и 2’дезокситимидинтрифосфатом соответственно. Мы часто сокращаем это обозначение до A, C, G и T. Обратите внимание, что две пары оснований имеют сходную структуру. А и G имеют два углеродно-азотных кольца и известны как пурины. Напротив, C и T имеют одно углеродно-азотное кольцо и принадлежат к классу молекул, называемых пиримидинами.
Взаимодействия водородных связей между основаниями позволяют двум нитям ДНК образовывать двойную спираль. Эти взаимодействия являются специфическими: пары оснований A с T и пары оснований C с G. Это происходит через водородные связи, которые показаны пунктирными линиями на рисунке выше. Если представить ДНК как винтовую лестницу, то ступенями были бы пары оснований. Ширина каждой «ступеньки» примерно одного размера, так как пара оснований всегда состоит из одного пиримидина и одного пурина. Нити ДНК идут антипараллельно или в противоположных направлениях: 5’-конец одной нити спаривается с 3’-концом другой. Это показано на рисунке ниже.
Эти взаимодействия являются специфическими: пары оснований A с T и пары оснований C с G. Это происходит через водородные связи, которые показаны пунктирными линиями на рисунке выше. Если представить ДНК как винтовую лестницу, то ступенями были бы пары оснований. Ширина каждой «ступеньки» примерно одного размера, так как пара оснований всегда состоит из одного пиримидина и одного пурина. Нити ДНК идут антипараллельно или в противоположных направлениях: 5’-конец одной нити спаривается с 3’-концом другой. Это показано на рисунке ниже.
Эта структура помещает неполярные основания ДНК в центр двухцепочечной молекулы, окруженной заряженными фосфатными группами. Это имеет два функциональных следствия. Во-первых, помните, что одноименные заряды отталкиваются друг от друга. Структура двойной спирали с отрицательно заряженными фосфатами на внешних краях позволяет фосфатам находиться как можно дальше друг от друга. Во-вторых, неполярные незаряженные основания спрятаны в центре спирали. Клеточная среда является водной и, следовательно, полярной, поэтому окружение неполярных оснований заряженными фосфатами максимизирует растворимость ДНК в физиологических условиях.
