Анализ спектра аминокислот и ацилкарнитинов, тандемная масс-спектрометрия, метод сухой капли крови (Analysis of amino acids and acylcarnitines, dried blood spots (MS/MS))
Метод определенияТандемная масс-спектрометрия с ионизацией в электроспрее.
Исследуемый материал Капиллярная кровь, собранная на специальную карточку-фильтр №903
Анализ спектра аминокислот и ацилкарнитинов методом тандемной масс-спектрометрии (ТМС)
Что такое нарушения метаболизма? Наследственные нарушения метаболизма или по-другому обмена веществ — это около 500 различных заболеваний, которые обусловлены нарушением работы особых биохимических катализаторов – ферментов. Ферменты обеспечивают процессы расщепления аминокислот, органических кислот, жирных кислот и других биомолекул. Многие ошибочно считают, что поскольку заболевания этой группы встречаются крайне редко, то и исключать их нужно в последнюю очередь. Однако по данным литературы*, наследственными нарушениями метаболизма страдает один из 3000 новорождённых!
Однако по данным литературы*, наследственными нарушениями метаболизма страдает один из 3000 новорождённых!
Особое место среди этих заболеваний занимают болезни, которые начинаются в раннем детском возрасте. Эти заболевания часто сочетаются с тяжёлой неонатальной патологией и/или протекают под маской таких состояний как сепсис, перинатальное поражение нервной системы, внутриутробная инфекция. Позднее выявление заболеваний этой группы может привести к тяжёлой инвалидности или даже летальному исходу. Установлено, что 5%** всех случаев «синдрома внезапной смерти младенцев» — следствие наследственных нарушений метаболизма. Однако некоторые из этих заболеваний эффективно лечатся при своевременной диагностике. Одним из современных методов диагностики нарушений метаболизма является тандемная масс-спектрометрия (ТМС). Этот метод позволяет определить в небольшом количестве биологического материала (капля высушенной крови) около 40 различных соединений, что позволяет с определённой вероятностью заподозрить наследственное заболевание.

Обращаем Ваше внимание на то, что из этого перечня в состав исследования «ПЯТОЧКА» входит только скрининг на фенилкетонурию (полный перечень выявляемых наследственных болезней обмена веществ при помощи скринига «ПЯТОЧКА» см. ниже по тексту).
На какие заболевания можно обследовать ребёнка дополнительно? Скрининга новорождённых, направленного на диагностику нарушений метаболизма методом ТМС, в России на настоящий момент не проводится. В России это исследование пока проводится по назначению врача при наличии подозрений на наследственные болезни обмена веществ, хотя многие из заболеваний этой группы проявляют себя не сразу после рождения, но при этом уже есть у новорождённого. Однако, уже упомянутым ранее методом тандемной масс-спектрометрии (ТМС) можно дополнительно обследовать новорождённого ребенка на исключение 37 различных наследственных заболеваний, которые относятся к нарушениям обмена аминокислот, органических кислот и дефектам ß -окисления жирных кислот. Аминоацидопатии Аминоацидопатии развиваются вследствие недостатка специфических ферментов, необходимых для метаболизма аминокислот.

- Болезнь с запахом кленового сиропа мочи (лейциноз).
- Цитрулинемия тип 1, неонатальная цитрулинемия.
- Аргининосукциновая ацидурия (АСА)/ недостаточность аргининосукцинат лиазы лиазы.
- Недостаточность орнитин транскарбамилазы.
- Недостаточность карбамилфосфат синтазы.
- Недостаточность N-ацетилглютамат синтазы.
- Некетотическая гиперглицинемия.
- Тирозинемия тип 1.
- Тирозинемия тип 2.
- Гомоцистинурия/недостаточность цистатионин бета-синтетазы.
- Фенилкетонурия.
- Аргининемия/недостаточность аргиназы.
- Пропионовая ацидемия (недостаточность пропионил КоА карбоксилазы).
- Метилмалоновая ацидемия.
- Изовалериановая ацидемия (недостаточность изовалерил КоА дегидрогеназы).
- Недостаточность 2-метилбутирил КоА дегидрогеназы.
- Недостаточность изобутирил КоА дегидрогеназы.
- Глутаровая ацидемия тип 1 (недостаточность глутарил КоА дегидрогеназы тип 1).
- Недостаточность 3-метилкротонил КоА карбоксилазы.
- Множественная карбоксилазная недостаточность.
- Недостаточность биотинидазы.
- Малоновая ацидемия (недостаточность малонил КоА декарбоксилазы).
- Недостаточность митохондриальной ацетоацетил КоА тиолазы.
- Недостаточность 2-метил-3-гидроксибутирил КоА дегидрогеназы.
- Недостаточность 3-гидрокси-3-метилглутарил КоА лиазы.
- Недостаточность 3-метилглутаконил КоА гидратазы.
- Недостаточность среднецепочечной ацил-КоА дегидрогеназы.
- Недостаточность очень длинноцепочечной ацил-КоА дегидрогеназы.

- Недостаточность короткоцепочечной ацил-КоА дегидрогеназы.
- Недостаточность длинноцепочечной 3-гидроксиацил-КоА дегидрогеназы (дефект трифункционального белка).
- Глутаровая ацидемия тип II (недостаточность глутарил КоА дегидрогеназы тип II), множественная недостаточность ацил-КоА дегидрогеназ.
- Нарушение транспорта карнитина.
- Недостаточность карнитин палмитоил трансферазы тип I.
- Недостаточность карнитин палмитоил трансферазы тип II.
- Недостаточность карнитин/ацилкарнитин транслоказы.
- Недостаточность 2,4-диеноил КоА редуктазы.
- Недостаточность среднецепочечной 3-кетоацил-КоА тиолазы.
- Недостаточность средне-/короткоцепочечной ацил-КоА дегидрогеназы.
4.2.2.1.1. Водорастворимые витамины \ КонсультантПлюс
4.2.2.1.1. Водорастворимые витамины.
Витамин C. Витамин C (формы и метаболиты аскорбиновой кислоты) участвует в окислительно-восстановительных реакциях, функционировании иммунной системы, способствует усвоению железа. Дефицит приводит к рыхлости и кровоточивости десен, носовым кровотечениям вследствие повышенной проницаемости и ломкости кровеносных капилляров. Среднее потребление варьирует в разных странах 70 — 170 мг/сут., в России — 55 — 70 мг/сут. Установленный уровень физиологической потребности в разных странах — 45 — 110 мг/сут. Верхний допустимый уровень потребления — 2 000 мг/сут.
Дефицит приводит к рыхлости и кровоточивости десен, носовым кровотечениям вследствие повышенной проницаемости и ломкости кровеносных капилляров. Среднее потребление варьирует в разных странах 70 — 170 мг/сут., в России — 55 — 70 мг/сут. Установленный уровень физиологической потребности в разных странах — 45 — 110 мг/сут. Верхний допустимый уровень потребления — 2 000 мг/сут.
Уточненная физиологическая потребность для взрослых — 90 мг/сут.
Физиологическая потребность для детей — от 30 до 90 мг/сут.
Витамин B1 (тиамин). Тиамин в форме образующегося из него тиаминдифосфата входит в состав важнейших ферментов углеводного и энергетического обмена, обеспечивающих организм энергией и пластическими веществами, а также метаболизм разветвленных аминокислот. Недостаток этого витамина ведет к серьезным нарушениям со стороны нервной, пищеварительной и сердечно-сосудистой систем. Среднее потребление варьирует в разных странах 1,1 — 2,3 мг/сут., в США — до 6,7 мг/сут., в России — 1,3 — 1,5 мг/сут. Установленный уровень потребности в разных странах — 0,9 — 2,0 мг/сут. Верхний допустимый уровень потребления не установлен.
Установленный уровень потребности в разных странах — 0,9 — 2,0 мг/сут. Верхний допустимый уровень потребления не установлен.
Уточненная физиологическая потребность для взрослых — 1,5 мг/сут.
Физиологическая потребность для детей — от 0,3 до 1,5 мг/сут.
Витамин B2 (рибофлавин). Рибофлавин в форме коферментов участвует в окислительно-восстановительных реакциях, способствует повышению восприимчивости цвета зрительным анализатором и темновой адаптации. Недостаточное потребление витамина B2 сопровождается нарушением состояния кожных покровов, слизистых оболочек, нарушением светового и сумеречного зрения. Среднее потребление в разных странах 1,5 — 7,0 мг/сут., в России — 1,0 — 1,3 мг/сут. Установленный уровень потребности в разных странах — 1,1 — 2,8 мг/сут. Верхний допустимый уровень потребления не установлен. При потреблении витамина B2 в размере 1,8 мг/сут. и более у подавляющего большинства обследованных лиц концентрация рибофлавина в сыворотке крови находится в пределах физиологической нормы.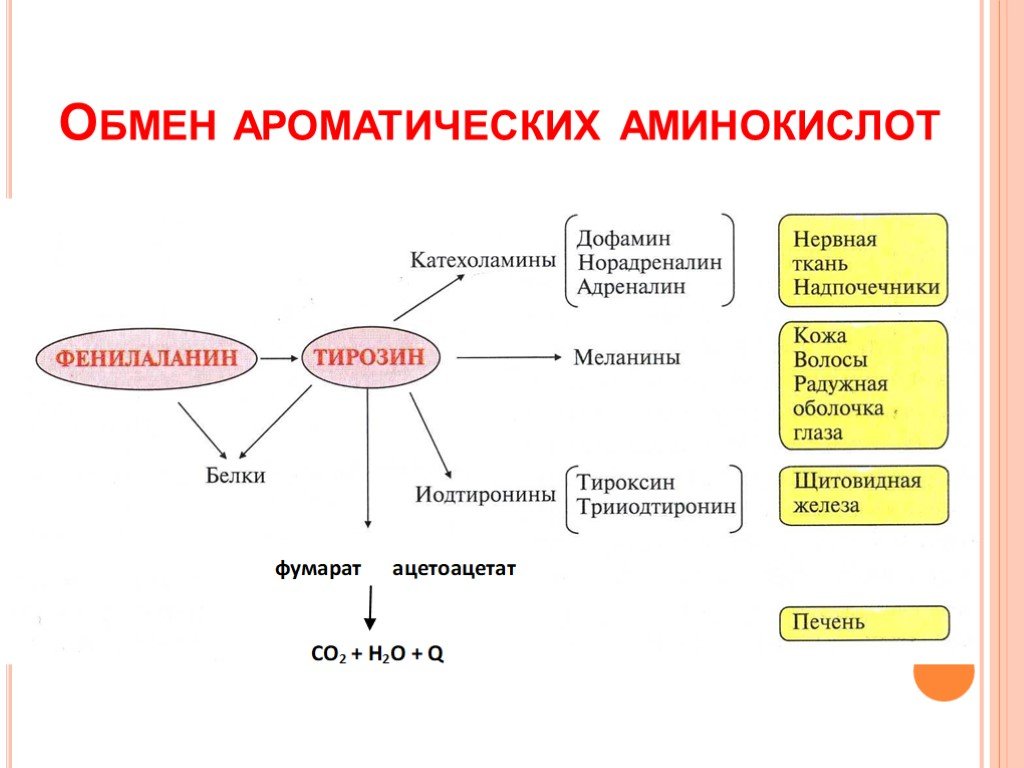
Уточненная физиологическая потребность для взрослых — 1,8 мг/сут.
Физиологическая потребность для детей — от 0,4 до 1,8 мг/сут.
Витамин B6 (пиридоксин). Пиридоксин в форме своих коферментов участвует в превращениях аминокислот, метаболизме триптофана, липидов и нуклеиновых кислот, участвует в поддержании иммунного ответа, процессах торможения и возбуждения в центральной нервной системе, способствует нормальному формированию эритроцитов, поддержанию нормального уровня гомоцистеина в крови. Недостаточное потребление витамина B6 сопровождается снижением аппетита, нарушением состояния кожных покровов, развитием гомоцистеинемии, анемии. Среднее потребление в разных странах 1,6 — 3,6 мг/сут., в Российской Федерации — 2,1 — 2,4 мг/сут. Недостаточная обеспеченность этим витамином обнаруживается у 50 — 70% населения Российской Федерации. Установленный уровень потребности в разных странах — 1,1 — 2,6 мг/сут. Верхний допустимый уровень потребления — 25,0 мг/сут.
Физиологическая потребность для взрослых — 2,0 мг/сут.
Физиологическая потребность для детей — от 0,4 до 2,0 мг/сут.
Ниацин. Ниацин в качестве кофермента участвует в окислительно-восстановительных реакциях энергетического метаболизма. Недостаточное потребление витамина сопровождается нарушением нормального состояния кожных покровов, желудочно-кишечного тракта и нервной системы. Среднее потребление в разных странах 12 — 40 мг/сут., в Российской Федерации — 13 — 15 мг/сут. Ниацин может синтезироваться из триптофана (из 60 мг триптофана образуется 1 мг ниацина). Установленный уровень потребности в разных странах — 11 — 25 мг/сут. Верхний допустимый уровень потребления ниацина — 60 мг/сут.
Физиологическая потребность для взрослых — 20 мг/сут.
Физиологическая потребность для детей — от 5 до 20 мг/сут.
Витамин B12. Витамин B12 играет важную роль в метаболизме и превращениях аминокислот. Фолат и витамин B12 являются взаимосвязанными витаминами, участвуют в кроветворении. Недостаток витамина B12 приводит к развитию частичной или вторичной недостаточности фолатов, а также анемии, лейкопении, тромбоцитопении. Среднее потребление в разных странах 4 — 17 мкг/сут., в Российской Федерации — около 3 мкг/сут. Установленный уровень потребности в разных странах — 1,4 — 3,0 мкг/сут. Верхний допустимый уровень потребления не установлен.
Недостаток витамина B12 приводит к развитию частичной или вторичной недостаточности фолатов, а также анемии, лейкопении, тромбоцитопении. Среднее потребление в разных странах 4 — 17 мкг/сут., в Российской Федерации — около 3 мкг/сут. Установленный уровень потребности в разных странах — 1,4 — 3,0 мкг/сут. Верхний допустимый уровень потребления не установлен.
Физиологическая потребность для взрослых — 3 мкг/сут.
Физиологическая потребность для детей — от 0,3 до 3,0 мкг/сут.
Фолаты. Фолаты в качестве кофермента участвуют в метаболизме нуклеиновых и аминокислот. Дефицит фолатов ведет к нарушению синтеза нуклеиновых кислот и белка, следствием чего является торможение роста и деления клеток, особенно в быстро пролифелирующих тканях: костный мозг, эпителий кишечника и др. Недостаточное потребление фолата во время беременности является одной из причин недоношенности, гипотрофии, врожденных уродств и нарушений развития ребенка. Показана выраженная связь между уровнем фолата, гомоцистеина и риском возникновения сердечно-сосудистых заболеваний. Среднее потребление в разных странах 210 — 400 мкг/сут. Установленный уровень потребности в разных странах — 150 — 400 мкг/сут. Верхний допустимый уровень потребления — 1 000 мкг/сут.
Среднее потребление в разных странах 210 — 400 мкг/сут. Установленный уровень потребности в разных странах — 150 — 400 мкг/сут. Верхний допустимый уровень потребления — 1 000 мкг/сут.
Уточненная физиологическая потребность для взрослых — 400 мкг/сут.
Физиологическая потребность для детей — от 50 до 400 мкг/сут.
Пантотеновая кислота. Пантотеновая кислота участвует в белковом, жировом, углеводном обмене, обмене холестерина, синтезе ряда гормонов, гемоглобина, способствует всасыванию аминокислот и сахаров в кишечнике, поддерживает функцию коры надпочечников. Недостаток пантотеновой кислоты может вести к поражению кожи и слизистых. Среднее потребление в разных странах 4,3 — 6,3 мг/сут. Установленный уровень потребности в разных странах — 4 — 12 мг/сут. Верхний допустимый уровень потребления не установлен.
Физиологическая потребность для взрослых — 5 мг/сут. (вводится впервые).
Физиологическая потребность для детей — от 1,0 до 5,0 мг/сут. (вводится впервые).
Биотин. Биотин участвует в синтезе жиров, гликогена, метаболизме аминокислот. Недостаточное потребление этого витамина может вести к нарушению нормального состояния кожных покровов. Среднее потребление в разных странах 20 — 53 мкг/сут. Установленный уровень потребности в разных странах — 15 — 100 мкг/сут. Верхний допустимый уровень потребления не установлен.
Биотин участвует в синтезе жиров, гликогена, метаболизме аминокислот. Недостаточное потребление этого витамина может вести к нарушению нормального состояния кожных покровов. Среднее потребление в разных странах 20 — 53 мкг/сут. Установленный уровень потребности в разных странах — 15 — 100 мкг/сут. Верхний допустимый уровень потребления не установлен.
Физиологическая потребность для взрослых — 50 мкг/сут. (вводится впервые).
Физиологическая потребность для детей — от 10 до 50 мкг/сут. (вводится впервые).
Метаболизм аминокислот
- Скачать PDF Копировать
Метаболизм аминокислот является важным процессом, происходящим в организме человека и помогающим в многочисленных биологических реакциях. В этой статье будет рассказано о роли глутамата, реакциях трансаминирования и различных типах аминокислот, таких как гликогенные, кетогенные и смешанные аминокислоты.
Реакции глутамата и трансаминирования
Четырьмя наиболее распространенными аминокислотами в организме человека являются глутамат, аспартат, аланин и глутамин, каждая из которых выполняет основные метаболические функции и роли в организме.
Глутамат имеет химическую структуру, аналогичную 2-оксоглутарату, который является промежуточным веществом в цикле Кребса в организме. Глутамат и 2-оксоглутарат находятся в равновесии и могут быть преобразованы трансаминазами или глутаматдегидрогеназой. Глутамат также может быть преобразован в глутамин, который является наиболее распространенной свободной аминокислотой в плазме крови человека и может быть переносчиком азота в организме.
Глутамат является наиболее распространенным и играет важную роль в функциях организма. Следует отметить, что когда дело доходит до метаболизма, глутамат играет центральную роль в расщеплении аминокислот. Например, пул глутамата необходим для выведения азота из пищевого белка из организма.
Трансаминирование — это реакция, включающая превращение аминокислоты в соответствующую кетокислоту. В этом типе реакции аминогруппа аминокислоты заменяется на другую кетокислоту, так что возникает новая пара аминокислоты и кетокислоты. В этом типе реакции нет чистой потери или увеличения азота. Реакции трансаминирования обратимы с константой равновесия, близкой к 1,9.0007
Введение в метаболизм аминокислот
Гликогенные, кетогенные и смешанные аминокислоты
Большинство углеродных скелетов аминокислот расщепляются до промежуточных продуктов цикла Кребса после трансаминирования. В результате они могут повышать уровень глюкозы в крови через глюконеогенный путь.
Related Stories
- Исследование показало, что современные противовирусные препараты сохраняют эффективность против недавно появившихся подвариантов Омикрона
- Исследование влияния добавок бета-аланина на работоспособность солдат
- Революционный дизайн белка: искусственный интеллект создает новые последовательности
Эти аминокислоты традиционно назывались «гликогенными» аминокислотами, поскольку было замечено, что они имеют тенденцию усугублять диабетическую глюкозурию по этой причине:
- Аланин: производится из и разлагается до пирувата
- Аргинин: производится из глутамата и расщепляется до него
- Аспарагин: производится из аспартата и расщепляется до него
- Аспартат: изготовлен из оксалоацетата и расщеплен до него
- Цистеин: незаменимая аминокислота, которая может быть получена из метионина и расщеплена до пирувата
- Глутамат: производится из оксоглутарата и расщепляется до него
- Глютамин: производится из глутамата и расщепляется до него
- Глицин: изготовлен из серина с множественными путями деградации
- Гистидин: незаменимая аминокислота, которая может расщепляться до глутамата
- Метионин: незаменимая аминокислота, которая может расщепляться до пропионил-КоА
- Пролин: производится из глутамата и расщепляется до него.
- Серин: производится из фосфоглицерата и расщепляется до пирувата
- Треонин: незаменимая аминокислота с неизвестными продуктами распада
- Валин: незаменимая аминокислота, которая может расщепляться до пропионил-КоА

«Кетогенные» аминокислоты имеют тенденцию усугублять диабетический кетоацидоз и обычно расщепляются до ацетоацетата или ацетил-КоА. Примером этого типа является лейцин, незаменимая аминокислота, которая может расщепляться до ацетил-КоА.
«Смешанные» аминокислоты могут расщепляться как до аминокислот цикла Кребса, так и до ацетил-КоА с характеристиками как гликогенных, так и кетогенных аминокислот.
- Изолейцин: незаменимая аминокислота, которая может расщепляться до ацетил-КоА и пропионил-КоА
- Лизин: незаменимая аминокислота с неизвестными продуктами распада
- Фенилаланин: незаменимая аминокислота, которая может расщепляться до тирозина
- Триптофан: незаменимая аминокислота с неизвестными продуктами распада
- Тирозин: незаменимая аминокислота, которая может быть получена из фенилаланина и расщеплена до фумарата и ацетоацетата.

Ссылки
- https://www.khanacademy.org/test-prep/mcat/biomolecules/fat-and-protein-metabolism/v/overview-of-amino-acid-metabolism
- https://www.slideshare.net/senchiy/амино-кислоты-метаболизм-new-12281450
- http://fblt.cz/en/skripta/ii-premena-latek-a-energie-v-bunce/12-metabolismus-aminokyselin/
- https://themedicalbiochemistrypage.org/amino-acid-metabolism.php
- http://www.bmb.leeds.ac.uk/illingworth/metabol/amino.htm
Последнее обновление: 26 февраля 2019 г.
- Скачать PDF Копировать
Используйте один из следующих форматов для ссылки на эту статью в своем эссе, статье или отчете:
APA
Smith, Yolanda. (2019, 26 февраля). Метаболизм аминокислот. Новости-Мед. Получено 4 июня 2023 г. с сайта https://www.news-medical.net/life-sciences/Amino-Acid-Metabolism.aspx.
MLA
Смит Иоланда.
 «Метаболизм аминокислот». Новости-Медицина . 4 июня 2023 г.
«Метаболизм аминокислот». Новости-Медицина . 4 июня 2023 г. Чикаго
Смит, Йоланда. «Метаболизм аминокислот». Новости-Мед. https://www.news-medical.net/life-sciences/Amino-Acid-Metabolism.aspx. (по состоянию на 04 июня 2023 г.).
Гарвард
Смит, Йоланда. 2019. Метаболизм аминокислот . News-Medical, просмотрено 4 июня 2023 г., https://www.news-medical.net/life-sciences/Amino-Acid-Metabolism.aspx.
Предлагаемая литература
8.1: Метаболизм аминокислот и специализированные продукты
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 37856
- Рене Дж.
 Леклер
Леклер - Virginia Tech Carilion School of Medicine via Virginia Tech Li Инициатива открытого образования braries
Кофакторы, необходимые для метаболизма аминокислот
Метаболизм многих аминокислот в значительной степени зависит от наличия кофакторов пиридоксальфосфата (витамин B 6 или PLP), тетрагидробиоптерина (BH 4 ) и тетрагидрофолата (TH 4 ). Важно признать, что недостатки этих кофакторов могут проявляться так же, как ферментативные дефициты определенных путей. Обратитесь к главе 2 для получения дополнительной информации о витамине B 6 и фолиевой кислоты.
Пиридоксальфосфат (B
6 или PLP)Во всех реакциях трансаминирования в качестве кофактора требуется PLP. Эти реакции необходимы для перемещения (или передачи) азота от аминокислоты к кетокислоте с образованием другой аминокислоты.
Тетрагидробиоптерин (BH
4 ) Это кофактор, синтезированный из GTP.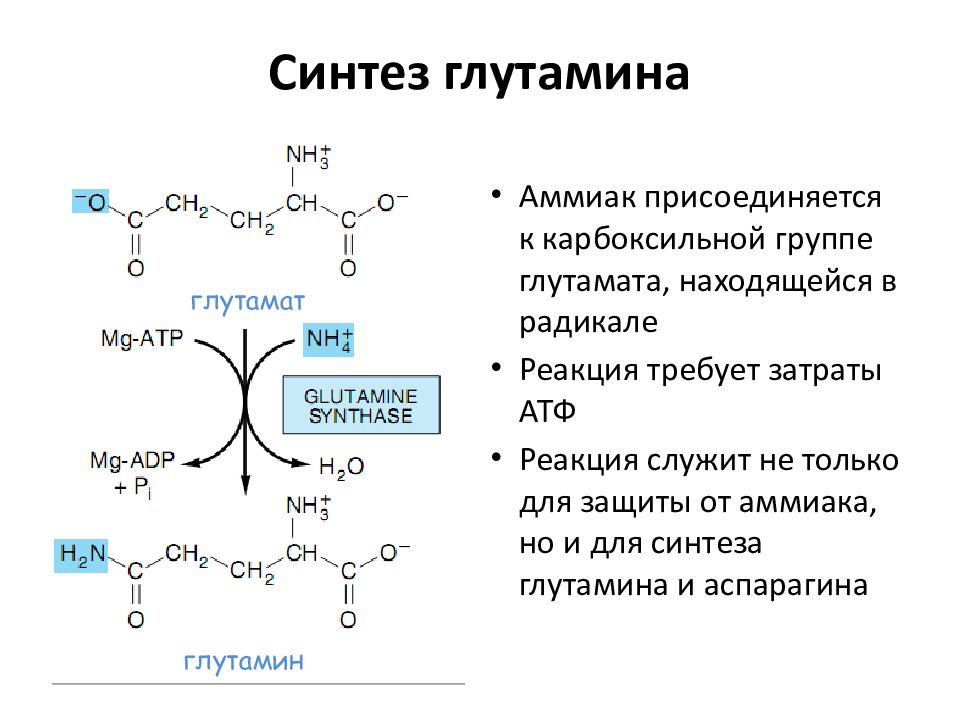 Он окисляется во время реакций гидроксилирования, особенно при превращении фенилаланина в тирозин. Дефицит ферментов, приводящий к снижению синтеза BH 4 может проявляться аналогично дефициту метаболизма фенилаланина.
Он окисляется во время реакций гидроксилирования, особенно при превращении фенилаланина в тирозин. Дефицит ферментов, приводящий к снижению синтеза BH 4 может проявляться аналогично дефициту метаболизма фенилаланина.
Тетрагидрофолат (FH
4 )Фолат может существовать во многих формах, и его часто называют тетрагидрофолатом. FH 4 часто встречается в различных формах с присоединенной одноуглеродной группой. Эти одноуглеродные группы, составляющие одноуглеродный пул, могут быть окислены или восстановлены. Одноуглеродные группы могут быть перенесены на другие соединения и играют существенную роль в синтезе глицина из серина, синтезе основания тимина (необходимого для синтеза ДНК), пуриновых оснований, необходимых для синтеза как ДНК, так и РНК, и переносе метильные группы в витамин \(B_{12}\).
Синтез специализированных продуктов
Ниже приведены некоторые ключевые аспекты метаболизма аминокислот.
Фенилаланин и тирозин
Фенилаланин является незаменимой аминокислотой, и гидроксилирование Phe фенилаланингидроксилазой (ПАУ) приводит к образованию тирозина (рис. 8.1). Для этого превращения требуется BH 4 , а дефицит кофактора или фермента PAH может привести к фенилкетонурии. Кроме того, неспособность синтезировать тирозин сделает эту аминокислоту условно незаменимой и потенциально негативно повлияет на синтез последующих соединений, показанных на рис. 8.1.
8.1). Для этого превращения требуется BH 4 , а дефицит кофактора или фермента PAH может привести к фенилкетонурии. Кроме того, неспособность синтезировать тирозин сделает эту аминокислоту условно незаменимой и потенциально негативно повлияет на синтез последующих соединений, показанных на рис. 8.1.
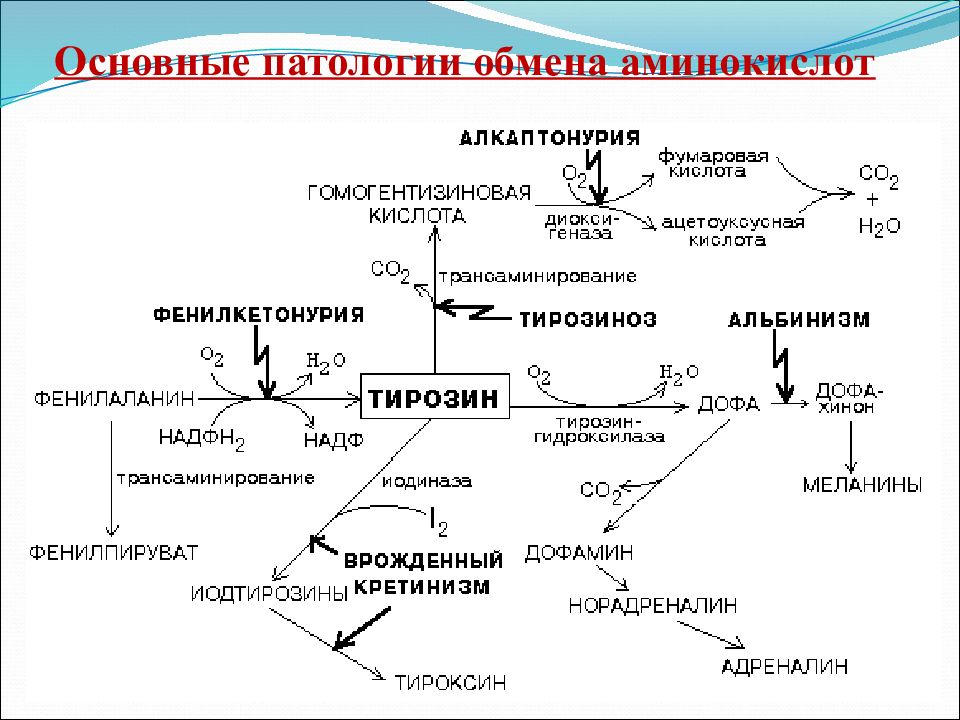
Тирозин может образовываться в результате метаболизма фенилаланина и необходим для производства меланина и катехоламинов. Дефицит может возникать в нескольких различных местах пути и приводить к альбинизму (тирозиназа), алкаптонурии (гомогентизатоксидаза) или тирозинемии, которые могут проявляться из-за дефицита нескольких ферментов пути (рис. 8.2).
Рисунок 8.2: Тирозин может образовываться в результате метаболизма фенилаланина и необходим для производства меланина и катехоламинов. Дефицит может возникать в нескольких различных местах пути и приводить к альбинизму, алкаптонурии или тирозинемии.
Фенилкетонурия
Фенилкетонурия (ФКУ) является одним из наиболее распространенных нарушений метаболизма аминокислот и наследуется по аутосомно-рецессивному типу. Симптомы нелеченной фенилкетонурии в первые месяцы жизни отсутствуют, поэтому скрининг новорожденных необходим для диагностики и начала лечения, что предотвращает разрушительные последствия детской гиперфенилаланинемии. Метод скрининга выявляет повышенные титры аминокислоты фенилаланина (Phe) в крови. Положительный результат теста (Phe более 150 мкмоль/л) побуждает врача начать использовать формулу с ограничением фенилаланина и требует подтверждающего количественного определения уровня Phe.
Глицин
Глицин является ключевым соединением, которое функционирует в качестве основного субстрата для различных путей, включая фолатный цикл, синтез нуклеотидов и синтез порфиринов (гема), глутатиона и креатина. Глицин может быть синтезирован на клеточном уровне из 3-фосфоглицерата, промежуточного продукта гликолиза.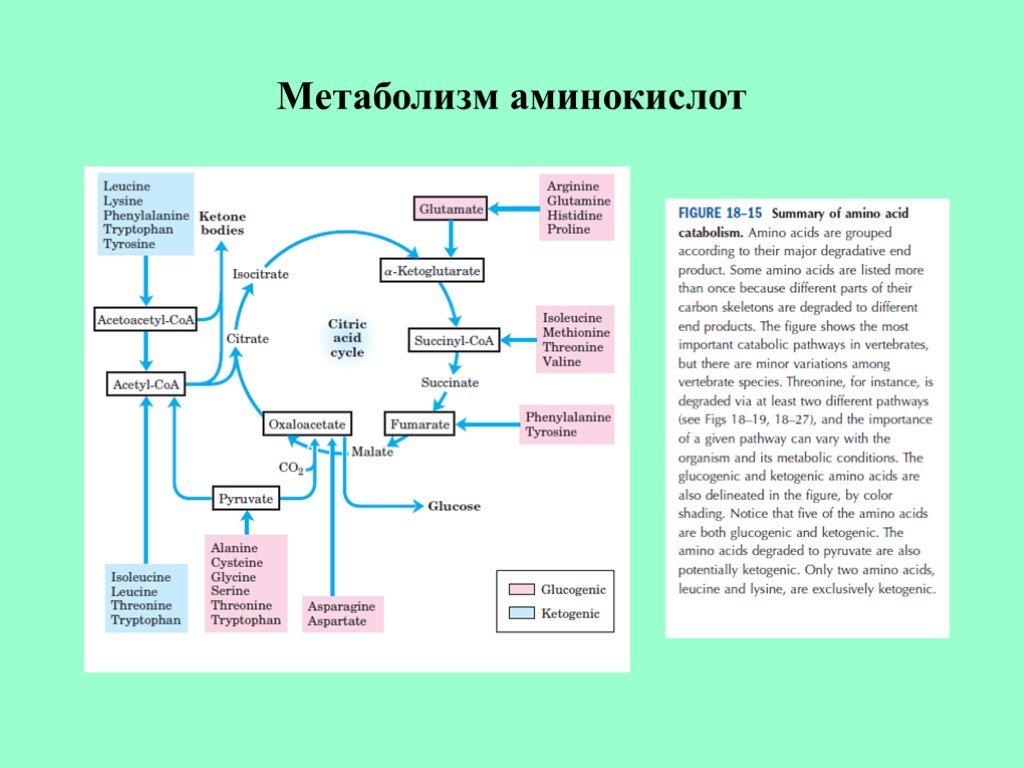
Аргинин
Аргинин — это заменимая аминокислота, поскольку она может быть получена в цикле мочевины. Дефицит цикла мочевины может привести к тому, что аргинин станет условно незаменимым. В этих случаях необходимы управление и добавки.
Триптофан
Триптофан является незаменимой аминокислотой, которая является одновременно кетогенной и глюкогенной, поскольку она может окисляться с образованием аланина и ацетил-КоА. Кольцевая структура также может быть использована для синтеза ниацина, что снижает потребность в этом витамине с пищей. Для метаболизма триптофана в серотонин (а затем и в мелатонин) требуется BH 4 . Недостатки здесь могут привести к дисбалансу этих нейротрансмиттеров (рис. 8.3).
Рисунок 8.3: Метаболизм триптофана в мелатонин.Глутамат
Глутамат играет ключевую роль в метаболизме аминокислот и обеспечивает субстраты для синтеза ГАМК и глутатиона (рис. 8.4). Кроме того, глутамат играет ключевую роль в перемещении азота в организме. Глутамат может быть дезаминирован глутаматдегидрогеназой с образованием \(\альфа\)-кетоглутарата. Он может входить непосредственно в цикл ТСА или подвергаться трансаминированию (рис. 8.4). Кроме того, глутамат можно использовать для фиксации или высвобождения аммония с образованием глютамина — одного из основных нетоксичных переносчиков аммиака.
Глутамат может быть дезаминирован глутаматдегидрогеназой с образованием \(\альфа\)-кетоглутарата. Он может входить непосредственно в цикл ТСА или подвергаться трансаминированию (рис. 8.4). Кроме того, глутамат можно использовать для фиксации или высвобождения аммония с образованием глютамина — одного из основных нетоксичных переносчиков аммиака.
Изолейцин, лейцин и валин (аминокислоты с разветвленной цепью)
Окисление этих аминокислот, в совокупности называемых аминокислотами с разветвленной цепью, происходит во всех тканях (кроме печени) и является основным источником топлива для скелетных мышц. Поскольку эти аминокислоты составляют примерно 25 % аминокислотного пула, они обеспечивают как энергию, так и доступный субстрат для восполнения цикла ТСА. Начальным этапом их метаболизма, как и всех аминокислот, является переаминирование с образованием кетокислоты. Затем эти соединения подвергаются окислительному декарбоксилированию с помощью многокомпонентного фермента, подобного комплексу пируватдегидрогеназы, с аналогичными требованиями к кофакторам (раздел 4. 1), а оставшиеся атомы углерода могут войти в цикл трикарбоновых кислот.
1), а оставшиеся атомы углерода могут войти в цикл трикарбоновых кислот.
Болезнь мочи, вызванная кленовым сиропом
Дефицит метаболизма аминокислот с разветвленной цепью может привести к диагнозу болезни мочи, вызванной кленовым сиропом (MSUD). С частотой 1 на 100 000 MSUD редко встречается даже среди врожденных нарушений метаболизма. Однако отчетливый сладкий запах, похожий на запах кленового сиропа, отличает это состояние как одно из наиболее узнаваемых нарушений обмена веществ. Это вызвано недостаточным окислительным декарбоксилированием \(\альфа\)-кетокислотных метаболитов лейцина, изолейцина и валина. У пораженных младенцев могут появиться симптомы в первые дни жизни, с плохим питанием, вялостью, судорогами и иногда комой. Более легкие формы MSUD могут проявиться в более позднем возрасте, с задержкой развития и умственной отсталостью. Болезнь мочи кленового сиропа в первую очередь лечится диетой, а также избеганием обстоятельств, усиливающих катаболизм, таких как высокая температура и обезвоживание. При возникновении метаболического криза необходимо экстренное лечение в стационаре для стабилизации состояния больного.
При возникновении метаболического криза необходимо экстренное лечение в стационаре для стабилизации состояния больного.
Метионин
Метионин является незаменимой аминокислотой со сложным метаболизмом, имеющим клиническое значение. Его метаболизм связан с фолатным циклом, реметилированием кобаламина и синтезом S-аденозилметионина (SAM). Дефицит ферментов или кофакторов может привести к повышению уровня гомоцистеина (гипергомоцистеинемия), что может иметь системные негативные последствия. Метионин, необходимый для синтеза SAM, может быть получен с пищей или произведен реметилированием гомоцистеина с использованием витамина \(B_{12}\).
Рисунок 8.5: Метаболизм аминокислот с разветвленной цепью. Дефицит дегидрогеназы кетокислот с разветвленной цепью (BCKAD) может привести к проявлению болезни мочи, вызванной кленовым сиропом. Сначала метионин конденсируется с АТФ с образованием SAM. SAM имеет заряженную метильную группу, которая может быть перенесена на множество различных молекул-акцепторов; этот шаг считается необратимым, поскольку количество высвобождаемой энергии является значительным. SAM используется многими биологическими путями для передачи метильных групп и пользуется постоянным спросом. После того, как SAM теряет свою метильную группу, образующееся соединение, S-аденозилгомоцистеин (SAH), гидролизуется до гомоцистеина и аденозина.
SAM используется многими биологическими путями для передачи метильных групп и пользуется постоянным спросом. После того, как SAM теряет свою метильную группу, образующееся соединение, S-аденозилгомоцистеин (SAH), гидролизуется до гомоцистеина и аденозина.
Гомоцистеин, полученный в результате этой реакции, может быть либо реметилирован в реакции с использованием как фолиевой кислоты, так и кобаламина для повторного синтеза метионина, либо может быть использован для синтеза цистеина (рис. 8.6).
Реметилирование гомоцистеина
Гомоцистеин может быть преобразован обратно в метионин с использованием как метил-ФГ 4 , так и витамина \(B_{12}\). (Это единственная реакция, в которой метил-FH 4 может отдавать метильную группу.) В этой реакции метильная группа из FH 4 переносится на кобаламин, связанный с гомоцистеинметилтрансферазой. Гомоцистеин получает метильную группу от этого заряженного кофактора кобаламина, а метионин регенерируется. Если гомоцистеинметилтрансфераза дефектна или уровень витамина \(B_{12}\) или FH 4 недостаточен, гомоцистеин будет накапливаться. Повышенный уровень гомоцистеина связывают с сердечно-сосудистыми и неврологическими заболеваниями. Следствием дефицита витамина \(B_{12}\) является накопление метил-ФГ 4 и снижение других производных фолиевой кислоты. Это известно как гипотеза метиловой ловушки; из-за дефицита \(B_{12}\) большая часть атомов углерода в пуле FH 4 захвачена в форме метил-FH 4 , которая является наиболее стабильной. Углерод не может быть высвобожден из фолата, потому что единственная реакция, в которой он участвует, не может происходить из-за дефицита \(B_{12}\). Это приводит к функциональному дефициту фолиевой кислоты, хотя общий уровень фолиевой кислоты в норме.
Если гомоцистеинметилтрансфераза дефектна или уровень витамина \(B_{12}\) или FH 4 недостаточен, гомоцистеин будет накапливаться. Повышенный уровень гомоцистеина связывают с сердечно-сосудистыми и неврологическими заболеваниями. Следствием дефицита витамина \(B_{12}\) является накопление метил-ФГ 4 и снижение других производных фолиевой кислоты. Это известно как гипотеза метиловой ловушки; из-за дефицита \(B_{12}\) большая часть атомов углерода в пуле FH 4 захвачена в форме метил-FH 4 , которая является наиболее стабильной. Углерод не может быть высвобожден из фолата, потому что единственная реакция, в которой он участвует, не может происходить из-за дефицита \(B_{12}\). Это приводит к функциональному дефициту фолиевой кислоты, хотя общий уровень фолиевой кислоты в норме.
Дефицит фолиевой кислоты (будь то функциональный или фактический) приводит к мегалобластной анемии, вызванной неспособностью предшественников клеток крови синтезировать ДНК и, следовательно, делиться. Это приводит к тому, что большие, частично реплицированные клетки выбрасываются в кровь, чтобы попытаться восполнить отмершие клетки. Дефицит фолиевой кислоты также был связан с увеличением частоты дефектов нервной трубки, таких как расщепление позвоночника, у матерей, которые забеременели при дефиците фолиевой кислоты.
Это приводит к тому, что большие, частично реплицированные клетки выбрасываются в кровь, чтобы попытаться восполнить отмершие клетки. Дефицит фолиевой кислоты также был связан с увеличением частоты дефектов нервной трубки, таких как расщепление позвоночника, у матерей, которые забеременели при дефиците фолиевой кислоты.
Путь транссульфурации
Дальнейший метаболизм гомоцистеина обеспечивает атом серы для синтеза цистеина. В этом двухстадийном процессе гомоцистеин сначала реагирует с серином с образованием цистатионина. Затем следует расщепление цистатионина с образованием цистеина и \(\альфа\)-кетобутирата. Первая реакция в этой последовательности, катализируемая цистатионин\(\бета\)-синтазой, ингибируется цистеином. Таким образом, метионин через гомоцистеин не используется для синтеза цистеина, если только уровень цистеина в организме не ниже, чем требуется для его метаболических функций. Таким образом, достаточное поступление цистеина с пищей может «уберечь» (или уменьшить) потребность в метионине с пищей (рис. 8.6).
8.6).
Последствия повышенного уровня гомоцистеина
Уровень гомоцистеина может накапливаться несколькими путями, связанными с метаболизмом как фолиевой кислоты, так и витамина \(B_{12}\). Поскольку SAM постоянно используется в качестве донора метила, это приводит к постоянному производству SAH. Следовательно, это приводит к постоянному производству гомоцистеина. Образовавшийся гомоцистеин может быть либо реметилирован в метионин, либо конденсирован с серином с образованием цистатионина. Основным путем метаболизма гомоцистеина является реметилирование с помощью N5-метил-Fh5, для которого требуется витамин \(B_{12}\). В печени также есть второй путь, по которому бетаин (продукт деградации холина) может отдавать метильную группу гомоцистеину с образованием метионина, но это второстепенный путь. Для превращения гомоцистеина в цистатионин требуется пиридоксальфосфат (PLP). Таким образом, если у человека дефицит витамина \(B_{12}\), подавляется превращение гомоцистеина в метионин основным путем. Это направляет гомоцистеин на производство цистатионина, который в конечном итоге производит цистеин. Гомоцистеин также накапливается в крови, если присутствует мутация в ферменте, превращающем N5,N10-метилен-Fh5 в N5-метил-Fh5. Когда это происходит, уровни N5-метил-Fh5 слишком низки, чтобы гомоцистеин мог превратиться в метионин. Утрата этого пути в сочетании с ингибированием цистеином образования цистатионина по принципу обратной связи также приводит к повышению уровня гомоцистеина в крови. Третьим способом повышения уровня гомоцистеина в сыворотке крови является мутация цистатиониновой \(\бета\)-синтазы или дефицит витамина \(B_{6}\), необходимого кофактора для этого фермента. Эти дефекты блокируют способность гомоцистеина к превращению в цистатионин, и гомоцистеин, который накапливается, не может быть полностью преобразован путем превращения в метионин.
Для превращения гомоцистеина в цистатионин требуется пиридоксальфосфат (PLP). Таким образом, если у человека дефицит витамина \(B_{12}\), подавляется превращение гомоцистеина в метионин основным путем. Это направляет гомоцистеин на производство цистатионина, который в конечном итоге производит цистеин. Гомоцистеин также накапливается в крови, если присутствует мутация в ферменте, превращающем N5,N10-метилен-Fh5 в N5-метил-Fh5. Когда это происходит, уровни N5-метил-Fh5 слишком низки, чтобы гомоцистеин мог превратиться в метионин. Утрата этого пути в сочетании с ингибированием цистеином образования цистатионина по принципу обратной связи также приводит к повышению уровня гомоцистеина в крови. Третьим способом повышения уровня гомоцистеина в сыворотке крови является мутация цистатиониновой \(\бета\)-синтазы или дефицит витамина \(B_{6}\), необходимого кофактора для этого фермента. Эти дефекты блокируют способность гомоцистеина к превращению в цистатионин, и гомоцистеин, который накапливается, не может быть полностью преобразован путем превращения в метионин. Таким образом, происходит накопление гомоцистеина.
Таким образом, происходит накопление гомоцистеина.
Ссылки и ресурсы
Текст
Ferrier, D. R., изд. Иллюстрированные обзоры Липпинкотта: биохимия , 7-е изд. Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins, 2017, Глава 20: Деградация и синтез аминокислот, Глава 21: Преобразование аминокислот в специализированные продукты.
Ле, Т. и В. Бхушан. Первая помощь для USMLE Step 1 , 29-е изд. Нью-Йорк: McGraw Hill Education, 2018, стр. 69, 83–85.
Либерман М. и А. Пит, ред. Базовая медицинская биохимия Маркса: клинический подход , 5-е изд. Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins, 2018, Глава 37: Синтез и деградация аминокислот, Глава 39: Тетрагидрофолат, витамин B12 и S-аденозилметионин.
Цифры
Серый, Сородич, Рисунок 8.1 Метаболизм фенилаланина требует Bh5, а также производит тирозин. Дефицит кофактора или фенилаланингидроксилазы может привести к фенилкетонурии. 2021. https://archive.org/details/8.1-new. CC BY 4.0 .
2021. https://archive.org/details/8.1-new. CC BY 4.0 .
Grey, Kindred, Рис. 8.2. Тирозин может образовываться в результате метаболизма фенилаланина и необходим для производства меланина и катехоламинов. Дефицит может возникать в нескольких различных местах пути и приводить к альбинизму, алкаптонурии или тирозинемии. 2021. https://archive.org/details/8.2_20210926. CC BY 4.0 .
Grey, Kindred, Рис. 8.3 Метаболизм триптофана в мелатонин. 2021. https://archive.org/details/8.3_20210926. CC BY 4.0 .
Grey, Kindred, Рис. 8.4 Метаболизм глутамата, взаимодействующий с транспортом азота и синтезом ГАМК. 2021. https://archive.org/details/8.4_20210926. CC BY 4.0 .
Grey, Kindred, Рис. 8.5 Метаболизм аминокислот с разветвленной цепью. Дефицит BCKAD может привести к болезни мочи, вызванной кленовым сиропом. 2021. https://archive.org/details/8.5_20210926. CC BY 4.0 .
Grey, Kindred, Рис. 8.6 Метаболизм метионина. Проиллюстрировано реметилирование и транссульфирование гомоцистеина. Дефицит кофакторов или ферментов может привести к повышению уровня гомоцистеина. 2021. https://archive.org/details/8.6_20210926. CC BY 4.0 .
8.6 Метаболизм метионина. Проиллюстрировано реметилирование и транссульфирование гомоцистеина. Дефицит кофакторов или ферментов может привести к повышению уровня гомоцистеина. 2021. https://archive.org/details/8.6_20210926. CC BY 4.0 .
Эта страница под заголовком 8.1: Метаболизм аминокислот и специализированные продукты распространяется по лицензии CC BY-NC-SA 4.0, автор, ремикширование и/или куратор Рене Дж. Леклер (Инициатива открытого образования Технических библиотек Вирджинии) .
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или страница
- Автор
- Рене Дж.

