Глюкоза в плазме
Глюкоза – это простой сахар, основной углеводород крови и главный источник энергии для всех клеток.
Синонимы русские
Анализ сахара в крови, глюкоза в крови, анализ глюкозы в крови натощак.
Синонимы английские
Blood sugar, fasting blood sugar, FBS, fasting blood glucose, FBG, fasting plasma glucose, blood glucose, urine glucose.
Метод исследования
Ферментативный УФ метод (гексокиназный).
Единицы измерения
Ммоль/л (миллимоль на литр), мг/дл (ммоль/л х 18,02 = мг/дл).
Какой биоматериал можно использовать для исследования?
Венозную, капиллярную кровь.
Как правильно подготовиться к исследованию?
- Не принимать пищу в течение 12 часов перед исследованием.
- Исключить физическое и эмоциональное перенапряжение за 30 минут до исследования.
- Не курить в течение 30 минут до исследования.


Общая информация об исследовании
Глюкоза – это простой сахар, служащий организму основным источником энергии. Употребляемые человеком углеводы расщепляются на глюкозу и другие простые сахара, которые усваиваются тонким кишечником и поступают в кровь.
Большинству клеток организма глюкоза требуется для выработки энергии. Мозгу же и нервным клеткам она нужна не только как источник энергии, но и как регулятор их деятельности, поскольку они могут функционировать, только если содержание глюкозы в крови достигает определенного уровня.
Организм может использовать глюкозу благодаря инсулину – гормону, вырабатываемому поджелудочной железой. Он регулирует движение глюкозы из крови в клетки организма, заставляя их накапливать избыток энергии в виде кратковременного резерва – гликогена либо в форме триглицеридов, откладывающихся в жировых клетках. Человек не может жить без глюкозы и без инсулина, содержание которых в крови должно быть сбалансировано.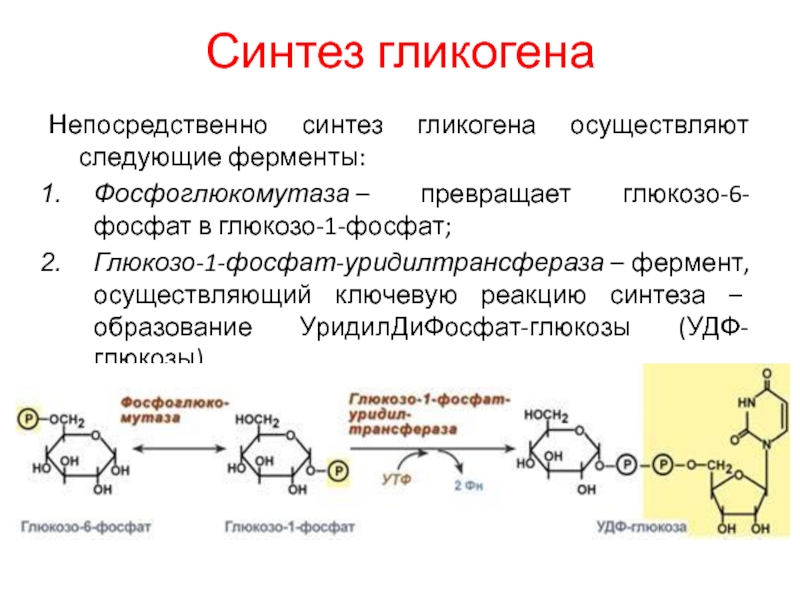
В норме содержание глюкозы в плазме крови слегка возрастает после еды, при этом секретирующийся инсулин понижает ее концентрацию. Уровень инсулина зависит от объема и состава принятой пищи. Если концентрация глюкозы в крови падает слишком низко, что может случаться после нескольких часов голодания либо после интенсивной физической работы, то выделяется глюкагон (еще один гормон поджелудочной железы), который заставляет клетки печени трансформировать гликоген обратно в глюкозу, тем самым повышая в крови ее содержание.
Поддержание нормы глюкозы в крови имеет крайне важное значение. Когда механизм обратной связи «глюкоза-инсулин» работает исправно, содержание глюкозы в крови остается достаточно стабильным. Если же этот баланс нарушается и уровень сахара в крови возрастает, то организм стремится восстановить его, во-первых, выработкой большего количества инсулина, а во-вторых, выведением глюкозы с мочой.
Крайние формы гипер- и гипогликемии (избытка и недостатка глюкозы) могут угрожать жизни больного, вызывая нарушение работы органов, повреждение мозга и кому. Хронически повышенное содержание глюкозы в крови может привести к повреждению почек, глаз, сердца, кровеносных сосудов и нервной системы. Хроническая гипогликемия опасна поражением мозга и нервной системы.
Хронически повышенное содержание глюкозы в крови может привести к повреждению почек, глаз, сердца, кровеносных сосудов и нервной системы. Хроническая гипогликемия опасна поражением мозга и нервной системы.
Иногда у женщин гипергликемия (гестационный диабет) возникает при беременности. Если ее не лечить, она может привести к тому, что у матери родится крупный ребенок с пониженным уровнем глюкозы в крови. Интересно, что женщина, страдающая от гипергликемии при беременности, после ее окончания не обязательно будет болеть диабетом.
Для чего используется исследование?
Уровень глюкозы важен при диагностике гипер- и гипогликемии и, соответственно, при диагностике сахарного диабета, а также для его последующего мониторинга. Анализ на сахар может быть сделан натощак (после 8-10 часов голодания), спонтанно (в любое время), после еды, а также может являться частью орального глюкозотолерантного теста (ГTT).
При выявлении диабета рекомендуется проводить анализ глюкозы в крови натощак либо глюкозотолерантный тест. Причем для окончательного подтверждения диагноза анализы должны проводиться двукратно в разное время.
Причем для окончательного подтверждения диагноза анализы должны проводиться двукратно в разное время.
Большинство беременных женщин проверяется на гестационный диабет (временную разновидность гипергликемии) между 24-й и 28-й неделями беременности.
Диабетики должны внимательно следить за уровнем глюкозы у себя в крови для корректировки приема таблетированных препаратов и выполнения инъекций инсулина. Обычно требуется по нескольку раз в день определять, насколько сильно отклоняется концентрация глюкозы от нормы.
Измерение уровня глюкозы в домашних условиях, как правило, осуществляется при помощи специального прибора – глюкометра, в который помещается тест-полоска с предварительно нанесенной каплей крови из пальца больного.
Когда назначается этот анализ?
- При профилактическом обследовании пациентов без подозрения на диабет, поскольку диабет – это заболевание, которое начинается с незначительных симптомов. Особенно важно следить за уровнем глюкозы в крови пациентам с генетической предрасположенностью к диабету, с повышенной массой тела и тем, кто старше 45 лет.

- При диагностике диабета у пациентов с симптомами гипер- или гипогликемии. Симптомы гипергликемии или повышенного сахара: повышенная жажда, усиленное мочеиспускание, утомляемость, неясность зрения, повышенная восприимчивость к инфекциям. Симптомы гипогликемии или пониженного сахара: потливость, повышенный аппетит, беспокойство, помутнение сознания, неясность зрения.
- При потере сознания или сильной слабости для выяснения, не вызваны ли они низким уровнем сахара в крови.
- Если у пациента зафиксировано преддиабетическое состояние (при котором содержание плазменной глюкозы выше нормы, но ниже, чем у больных диабетом), анализ проводится через регулярные интервалы.
- Лицам, у которых диагностирован сахарный диабет, тест на глюкозу в крови назначают совместно с анализом на гликированный гемоглобин (А1с), чтобы проследить изменение содержания глюкозы в крови за длительный промежуток времени.
- В некоторых случаях анализ на глюкозу в плазме крови может проводиться совместно с анализом на инсулин и С-пептид для мониторинга выработки инсулина.

- Беременные обычно проверяются на гестационный диабет в конце срока. Если у женщины был обнаружен гестационный диабет до этого, то она сдает анализ на глюкозу в течение всей беременности, а также после родов.
Что означают результаты?
Референсные значения (Норма глюкозы в крови)
|
Возраст |
Референсные значения |
|
меньше 14 лет |
3,3 – 5,6 ммоль/л |
|
дети старше 14 лет, мужчины, небеременные женщины |
4,1 – 6,1 ммоль/л |
|
беременные женщины |
4,1 – 5,1 ммоль/л |
Другие причины повышенного уровня глюкозы:
- акромегалия,
- сильный стресс (реакция на травму, сердечный приступ, инсульт),
- хроническая почечная недостаточность,
- синдром гиперкортицизма (Иценко – Кушинга),
- прием таких лекарств, как кортикостероиды, антидепрессанты трициклического ряда, диуретики, эпинефрины, эстрогены, литий, дифенин (дилантин), салицилаты,
- избыточное потребление высокоуглеводной пищи,
- гипертиреоз,
- рак поджелудочной железы,
- панкреатит.
Причины пониженного уровня глюкозы в крови:
- недостаточность надпочечников,
- злоупотребление алкоголем,
- принятие таких препаратов, как ацетаминофен и анаболические стероиды,
- болезни печени,
- гипопитуитаризм,
- гипотиреоз,
- передозировка инсулина,
- инсулиномы,
- голодание.
Важные замечания
- Гипогликемия характеризуется падением содержания в плазме крови глюкозы до уровня, при котором наступают расстройства нервной системы (потливость, дрожь, чувство голода, беспокойство), затем подвергается воздействию и головной мозг (помутнение сознания, галлюцинации, неясность зрения, иногда кома и даже смерть).
- Для уверенной постановки диагноза «гипогликемия» требуется подтверждение «триады Вайпла»:
- уровень глюкозы ниже порога 40 мг/дл (2,2 ммоль/л),
- симптомы гипогликемии,
- исчезновение симптомов, когда содержание глюкозы в крови возвращается к норме.

- Первичная гипогликемия является редкой формой и обычно диагностируется в детстве.
- Признаки гипогликемии иногда проявляются у пациентов, у которых уровень сахара в крови не понижен. В таком случае изменение питания, например более частое употребление еды в течение дня мелкими порциями и выбор сложных углеводов, может быть достаточным, чтобы избавиться от симптомов.
Также рекомендуется
- Гликированный гемоглобин (HbA1c)
- С-пептид в сыворотке
- Инсулин
- Глюкоза в моче
- Общий анализ мочи с микроскопией
- Альбумин в моче (микроальбуминурия) или Альбумин-креатининовое соотношение (альбуминурия в разовой порции мочи)
Кто назначает исследование?
Терапевт, эндокринолог, педиатр, врач общей практики, гастроэнтеролог.
Литература
- Федеральные клинические рекомендации (протоколы) по ведению детей с эндокринными заболеваниями.

- Клинические рекомендации «Алгоритмы специализированной медицинской помощи больным сахарным диабетом». Под редакцией И.И. Дедова, М.В. Шестаковой, А.Ю. Майорова 8-й выпуск, М., 2017.
- Definition and diagnosis of diabetes mellitus and intermediate hyperglycemia. Report of a WHO/IDF ConsultatIon. 2006.
- Рекомендации по диабету, предиабету и сердечно-сосудистым заболеваниям. EASD/ESC, Российский кардиологический журнал 2014; № 3(107):7-61.
- Гестационный сахарный диабет: диагностика, лечение, послеродовое наблюдение. Клинические рекомендации (протокол) МЗ РФ. М., 2014.
КАТАБОЛИЗМ И БИОСИНТЕЗ ГЛИКОГЕНА — TDMUV
КАТАБОЛИЗМ И БИОСИНТЕЗ ГЛИКОГЕНА. РЕГУЛЯЦИЯ ОБМЕНА ГЛИКОГЕНА.
БИОСИНТЕЗ ГЛЮКОЗЫ — ГЛЮКОНЕОГЕНЕЗ. МЕХАНИЗМЫ ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ И ПАТОЛОГИЯ ОБМЕНА УГЛЕВОДОВ.
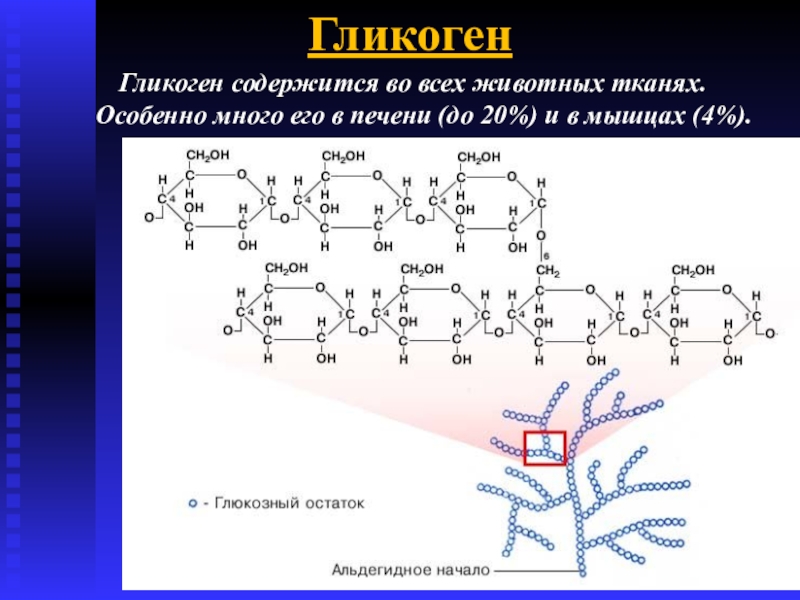
Гликоген –
главная форма запасания углеводов
у животных и человека.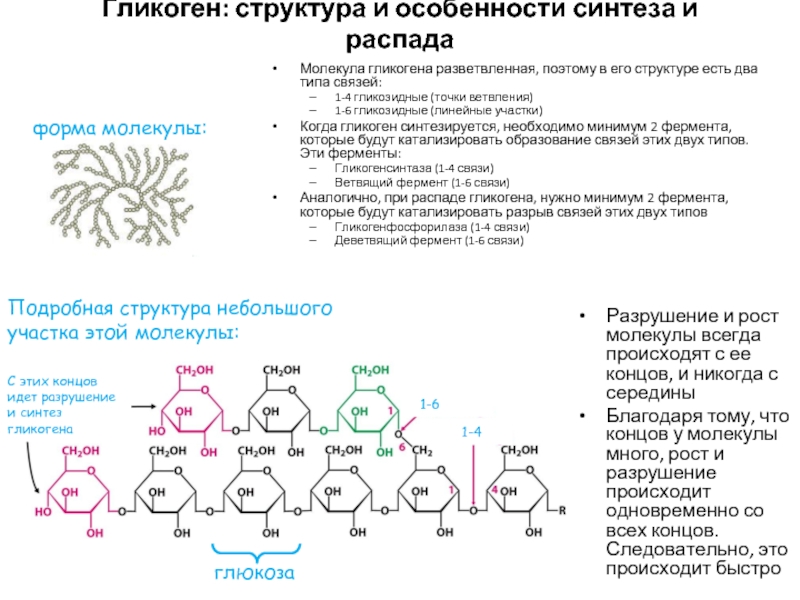 Накапливается гликоген главным образом в
печени (до 6% от массы печени) и в скелетных мышцах, где его содержание редко
превышает 1%. Запасы гликогена
в скелетных мышцах ввиду значительно большей массы последних превышают его
запасы в печени. Гликоген присутствует в
цитозоле в форме гранул диаметром от 10 до 40 нм. На электронных микрофотографиях
гликогеновые гранулы выглядят плотными. Установлено, что эти гранулы, кроме гликогена, содержат ферменты, катализирующие
синтез и распад гликогена.
Однако гликогеновые гранулы отличаются от мульти-ферментных комплексов
(например, от пируватдегидрогеназного комплекса). Степень структурной
организации гликогеновых гранул ниже, чем в мультиферментных комплексах.
Следует подчеркнуть, что синтез и распад гликогена в клетке осуществляются
разными метаболическими путями. В частности, считалось, что гликогенфосфорилаза
(фосфорилаза а) катализирует как распад гликогена, так и его
синтез, потому что в опытах in vitro было показано, что гликогенфосфорилазная реакция
обратима.
Накапливается гликоген главным образом в
печени (до 6% от массы печени) и в скелетных мышцах, где его содержание редко
превышает 1%. Запасы гликогена
в скелетных мышцах ввиду значительно большей массы последних превышают его
запасы в печени. Гликоген присутствует в
цитозоле в форме гранул диаметром от 10 до 40 нм. На электронных микрофотографиях
гликогеновые гранулы выглядят плотными. Установлено, что эти гранулы, кроме гликогена, содержат ферменты, катализирующие
синтез и распад гликогена.
Однако гликогеновые гранулы отличаются от мульти-ферментных комплексов
(например, от пируватдегидрогеназного комплекса). Степень структурной
организации гликогеновых гранул ниже, чем в мультиферментных комплексах.
Следует подчеркнуть, что синтез и распад гликогена в клетке осуществляются
разными метаболическими путями. В частности, считалось, что гликогенфосфорилаза
(фосфорилаза а) катализирует как распад гликогена, так и его
синтез, потому что в опытах in vitro было показано, что гликогенфосфорилазная реакция
обратима.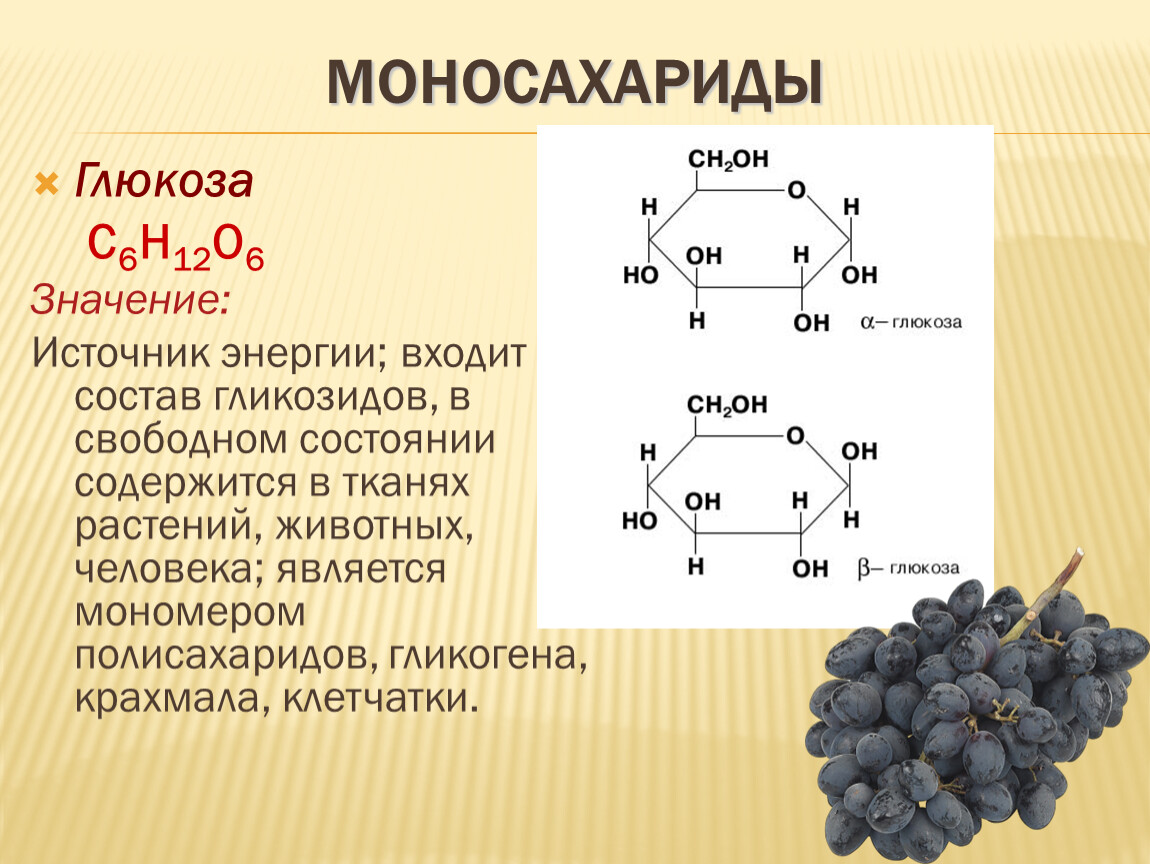 Однако в дальнейшем было установлено, что в клетке (in vivo) фосфорилаза а катализирует только распад гликогена, синтез гликогена осуществляется
при участии совершенно другого фермента. Оба эти процесса
(синтез и распад гликогена) регулируют содержание глюкозы в крови и создают
резерв глюкозы
для интенсивной мышечной работы.
Однако в дальнейшем было установлено, что в клетке (in vivo) фосфорилаза а катализирует только распад гликогена, синтез гликогена осуществляется
при участии совершенно другого фермента. Оба эти процесса
(синтез и распад гликогена) регулируют содержание глюкозы в крови и создают
резерв глюкозы
для интенсивной мышечной работы.
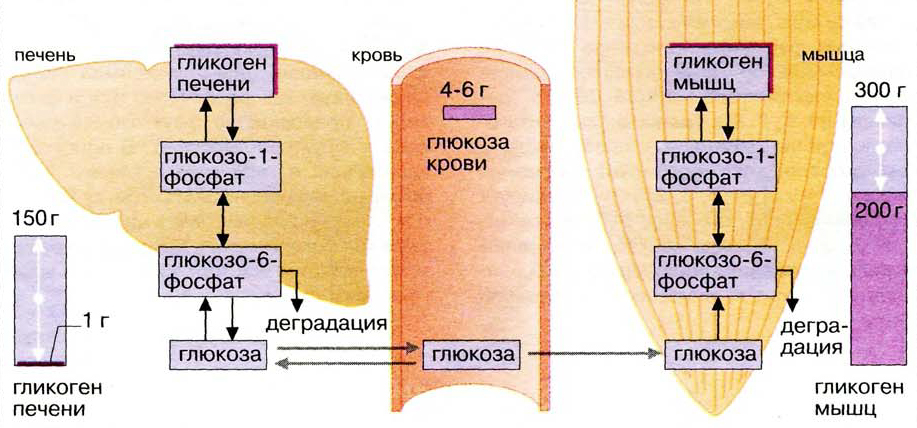
Депонирование и распад гликогена
Гликоген — основная форма депонирования
глюкозы в клетках животных. У растений эту же функцию выполняет крахмал. В
структурном отношении гликоген, как и крахмал, представляет собой разветвленный
полимер из глюкозы: Однако гликоген более разветвлен и компактен. Ветвление
обеспечивает быстрое освобождение при распаде гликогена большого количества
концевых мономеров. Синтез и распад гликогена не являются обращением друг в
друга, эти процессы происходят разными путями: Гликоген синтезируется в
период пищеварения (в течение 1-2 часов после приема углеводной пищи). Гликогенез особенно интенсивно протекает в печени и скелетных мышцах. В
начальных реакциях образуется UDF-глюкоза (реакция 3), которая является
активированной формой глюкозы, непосредственно включающейся в реакцию
полимеризации (реакция 4). Эта последняя реакция катализируется гликогенсинтазой,
которая присоединяет глюкозу к олигосахариду или к уже имеющейся в клетке
молекуле гликогена, наращивая цепь новыми мономерами. Для подготовки и
включения в растущую полисахаридную цепь требуется энергия 1 моль АТР и 1 моль
UTP. Ветвление полисахаридной цепи происходит при участии
фермента амило- a -1,4— a -1,6-гликозил-трансферазы путем
разрыва одной a -1,4-связи и переноса олигосахаридного остатка от конца
растущей цепи к ее середине с образованием в этом месте a -1,6-гликозидной
связи . Молекула гликогена содержит до 1 млн. остатков
глюкозы, следовательно, на синтез расходуется значительное количество энергии.
Гликогенез особенно интенсивно протекает в печени и скелетных мышцах. В
начальных реакциях образуется UDF-глюкоза (реакция 3), которая является
активированной формой глюкозы, непосредственно включающейся в реакцию
полимеризации (реакция 4). Эта последняя реакция катализируется гликогенсинтазой,
которая присоединяет глюкозу к олигосахариду или к уже имеющейся в клетке
молекуле гликогена, наращивая цепь новыми мономерами. Для подготовки и
включения в растущую полисахаридную цепь требуется энергия 1 моль АТР и 1 моль
UTP. Ветвление полисахаридной цепи происходит при участии
фермента амило- a -1,4— a -1,6-гликозил-трансферазы путем
разрыва одной a -1,4-связи и переноса олигосахаридного остатка от конца
растущей цепи к ее середине с образованием в этом месте a -1,6-гликозидной
связи . Молекула гликогена содержит до 1 млн. остатков
глюкозы, следовательно, на синтез расходуется значительное количество энергии.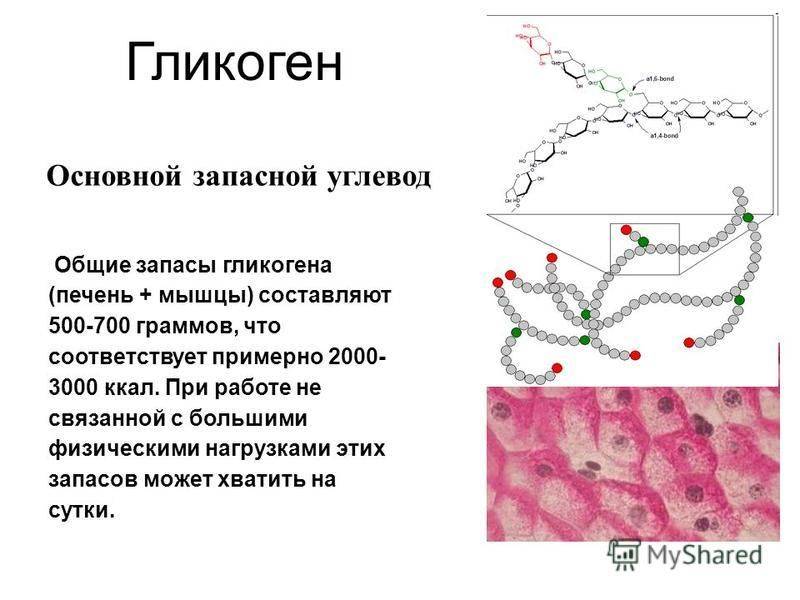 Необходимость превращения глюкозы в гликоген связана с тем, что накопление
значительного количества глюкозы в клетке привело бы к повышению осмотического
давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген
содержится в клетке в виде гранул, и мало растворим. Распад
гликогена — гликогенолиз — происходит в период между приемами пищи.
Освобождение глюкозы в виде глюкозо-1-фосфата ( реакция 5 ) происходит в
результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет
концевые остатки один за другим, укорачивая цепи гликогена. Однако этот фермент
расщепляет только a -1,4 гликозидные связи. Связи в точке ветвления
гидролизуются ферментом амило- a
-1,6-гликозидазой , который отщепляет мономер глюкозы в
свободном виде.
Необходимость превращения глюкозы в гликоген связана с тем, что накопление
значительного количества глюкозы в клетке привело бы к повышению осмотического
давления, так как глюкоза хорошо растворимое вещество. Напротив, гликоген
содержится в клетке в виде гранул, и мало растворим. Распад
гликогена — гликогенолиз — происходит в период между приемами пищи.
Освобождение глюкозы в виде глюкозо-1-фосфата ( реакция 5 ) происходит в
результате фосфоролиза, катализируемого фосфорилазой. Фермент отщепляет
концевые остатки один за другим, укорачивая цепи гликогена. Однако этот фермент
расщепляет только a -1,4 гликозидные связи. Связи в точке ветвления
гидролизуются ферментом амило- a
-1,6-гликозидазой , который отщепляет мономер глюкозы в
свободном виде.
Особенности метаболизма гликогена в печени и мышцах
Включение глюкозы в метаболизм начинается с образования
фосфоэфира — глюкозо-6-фосфата. В клетках мышц и других органах
эту реакцию катализирует фермент гексокиназа, его Км менее 0,1 ммоль/л. В клетках печени эту же
реакцию катализирует глюкокиназа, значение Км которой примерно 10
ммоль/л. Это значит, что насыщение глюкокиназы происходит только при высокой
концентрации глюкозы. Различия в свойствах ферментов объясняют, почему в период
пищеварения глюкоза задерживается в основном в печени. Глюкокиназа при высокой
концентрации глюкозы в этот период максимально активна.
Напротив, гексокиназа, обладая большим сродством к глюкозе, способна
выхватывать ее из общего кровотока, где концентрация глюкозы ниже. Физиологическое
значение гликогенолиза в печени и в мышцах различно. Мышечный гликоген
является источником глюкозы для самой клетки. Гликоген печени используется
главным образом для поддержания физиологической концентрации глюкозы в крови.
Различия обусловлены тем, что в клетке печени присутствует фермент глюкозо-6-фосфатаза,
катализирующая отщепление фосфатной группы и образование свободной глюкозы,
после чего глюкоза поступает в кровоток.
В клетках мышц и других органах
эту реакцию катализирует фермент гексокиназа, его Км менее 0,1 ммоль/л. В клетках печени эту же
реакцию катализирует глюкокиназа, значение Км которой примерно 10
ммоль/л. Это значит, что насыщение глюкокиназы происходит только при высокой
концентрации глюкозы. Различия в свойствах ферментов объясняют, почему в период
пищеварения глюкоза задерживается в основном в печени. Глюкокиназа при высокой
концентрации глюкозы в этот период максимально активна.
Напротив, гексокиназа, обладая большим сродством к глюкозе, способна
выхватывать ее из общего кровотока, где концентрация глюкозы ниже. Физиологическое
значение гликогенолиза в печени и в мышцах различно. Мышечный гликоген
является источником глюкозы для самой клетки. Гликоген печени используется
главным образом для поддержания физиологической концентрации глюкозы в крови.
Различия обусловлены тем, что в клетке печени присутствует фермент глюкозо-6-фосфатаза,
катализирующая отщепление фосфатной группы и образование свободной глюкозы,
после чего глюкоза поступает в кровоток.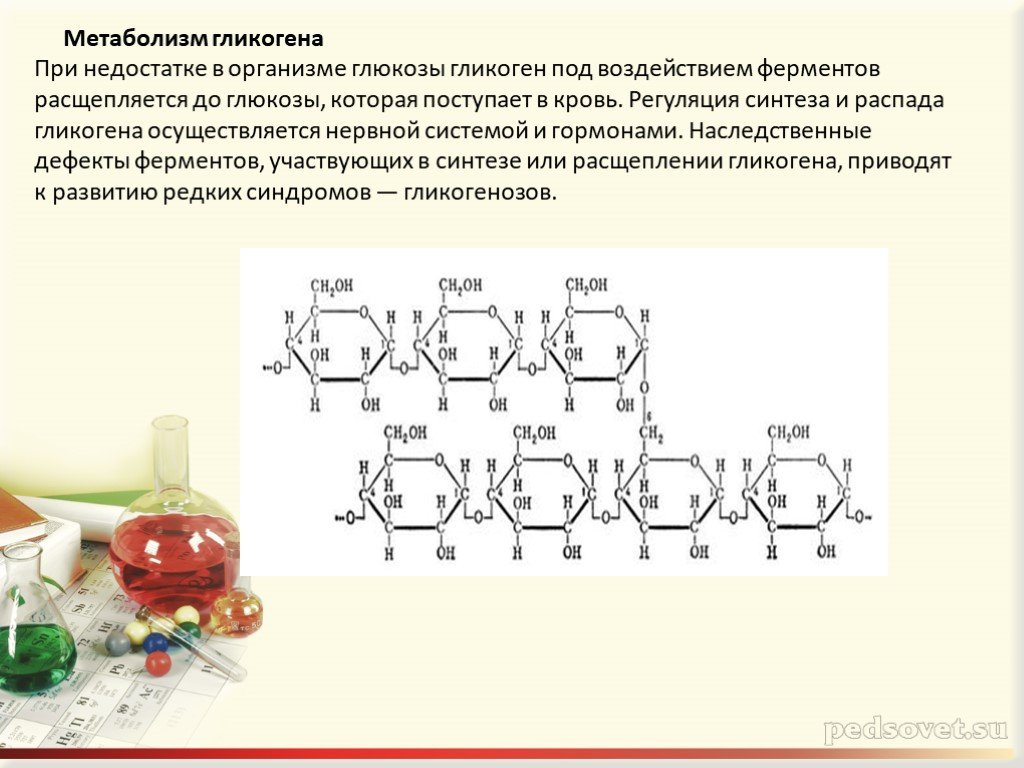 В клетках мышц нет этого
фермента, и распад гликогена идет только до образования
глюкозо-6-фосфата, который затем используется в клетке.
В клетках мышц нет этого
фермента, и распад гликогена идет только до образования
глюкозо-6-фосфата, который затем используется в клетке.
Биосинтез глюкозы — глюконеогенез
Глюконеогенез — это синтез глюкозы из неуглеводных
предшественников. У млекопитающих эту функцию выполняет
в основном печень, в меньшей мере — почки и клетки
слизистой кишечника. Запасов гликогена в организме
достаточно для удовлетворения потребностей в глюкозе в период между приемами
пищи. При углеводном или полном голодании, а также в условиях длительной физической
работы концентрация глюкозы в крови поддерживается за счет глюконеогенеза. В
этот процесс могут быть вовлечены вещества, которые способны превратиться в
пируват или любой другой метаболит глюконеогенеза. Причем, использование
первичных субстратов в глюконеогенезе происходит в различных физиологических
состояниях. Так, в условиях голодания часть тканевых белков распадается до
аминокислот, которые затем используются в глюконеогенезе. При распаде жиров
образуется глицерин, который через диоксиацетонфосфат включается в
глюконеогенез. Лактат, образующийся при интенсивной
физической работе в мышцах, затем в печени превращается в глюкозу.
Следовательно, физиологическая роль глюконеогенеза из лактата и из аминокислот
и глицерина различна. Синтез глюкозы из пирувата протекает, как и при
гликолизе, но в обратном направлении: Семь реакций гликолиза легко обратимы и
используются в глюконеогенезе. Но три киназные реакции необратимы и должны
шунтироваться. Так, фруктозо-1,6-дифосфат и глюкозо-6-фосфат дефосфорилируются
специфическими фосфатазами, а пируват фосфорилируется до образования
фосфоенолпирувата посредством двух промежуточных стадий через оксалоацетат.
Образование оксалоацетата катализируется пируваткарбоксилазой. Этот фермент
содержит в качестве кофермента биотин.
Так, в условиях голодания часть тканевых белков распадается до
аминокислот, которые затем используются в глюконеогенезе. При распаде жиров
образуется глицерин, который через диоксиацетонфосфат включается в
глюконеогенез. Лактат, образующийся при интенсивной
физической работе в мышцах, затем в печени превращается в глюкозу.
Следовательно, физиологическая роль глюконеогенеза из лактата и из аминокислот
и глицерина различна. Синтез глюкозы из пирувата протекает, как и при
гликолизе, но в обратном направлении: Семь реакций гликолиза легко обратимы и
используются в глюконеогенезе. Но три киназные реакции необратимы и должны
шунтироваться. Так, фруктозо-1,6-дифосфат и глюкозо-6-фосфат дефосфорилируются
специфическими фосфатазами, а пируват фосфорилируется до образования
фосфоенолпирувата посредством двух промежуточных стадий через оксалоацетат.
Образование оксалоацетата катализируется пируваткарбоксилазой. Этот фермент
содержит в качестве кофермента биотин. Оксалоацетат образуется в митохондриях,
транспортируется в цитозоль и включается в глюконеогенез. Следует обратить
внимание на то, что каждая из необратимых реакций гликолиза вместе с
соответствующей ей необратимой реакцией глюконеогенеза составляют цикл,
называемый субстратным: Таких циклов существует три — соответственно трем
необратимым реакциям. Результатом одновременного протекания реакций субстратных
циклов будет расходование энергии. Субстратные циклы могут протекать в условиях
нормального обмена веществ в печени и имеют вполне определенное биологическое
значение. Кроме того, эти циклы служат точками приложения регуляторных
механизмов, в результате чего изменяется поток метаболитов либо по пути
распада глюкозы, либо по пути ее синтеза. Суммарное уравнение глюконеогенеза из
пирувата:
Оксалоацетат образуется в митохондриях,
транспортируется в цитозоль и включается в глюконеогенез. Следует обратить
внимание на то, что каждая из необратимых реакций гликолиза вместе с
соответствующей ей необратимой реакцией глюконеогенеза составляют цикл,
называемый субстратным: Таких циклов существует три — соответственно трем
необратимым реакциям. Результатом одновременного протекания реакций субстратных
циклов будет расходование энергии. Субстратные циклы могут протекать в условиях
нормального обмена веществ в печени и имеют вполне определенное биологическое
значение. Кроме того, эти циклы служат точками приложения регуляторных
механизмов, в результате чего изменяется поток метаболитов либо по пути
распада глюкозы, либо по пути ее синтеза. Суммарное уравнение глюконеогенеза из
пирувата:
2 пируват + 4 ATP + 2 GTP + 2(NADH) + 4 Н2О ® Глюкоза + 4 ADP + 2 GDP + 2 NAD+ + 6 Н3РО4
За сутки в организме человека может синтезироваться до 80 г глюкозы. На синтез 1 моль
глюкозы из пирувата расходуется 6 макроэргических связей (4 ATP и 2 GTP).
На синтез 1 моль
глюкозы из пирувата расходуется 6 макроэргических связей (4 ATP и 2 GTP).
Глюкозо-лактатный цикл (цикл Кори)
Начинается с образования лактата в мышцах в результате анаэробного гликолиза (особенно в белых мышечных волокнах, которые бедны митохондриями по сравнению с красными). Лактат переносится кровью в печень, где в процессе глюконеогенеза превращается в глюкозу, которая затем с током крови может возвращаться в работающую мышцу. Итак печень снабжает мышцу глюкозой и, следовательно, энергией для сокращений. В печени часть лактата может окисляться до СО2 и Н2О, превращаясь в пируват и — далее в общих путях катаболизма.
Пентозофосфатный путь в метаболизме глюкозы
Это путь превращения глюкозы в пентозы. В
пентозофосфатном пути превращения глюкозы можно выделить две части: А -
окислительный путь и Б -
неокислительный путь синтеза пентоз. Коферментом дегидрогеназ является
NADP+, который восстанавливается в NADPH и используется клетками в
реакции восстановления и гидроксилирования. Кроме того,
пентозофосфатный путь (окислительный и неокислительный) поставляет клетке пентозофосфаты,
необходимые для синтеза нуклеиновых кислот и
коферментов (NAD, FAD, СоА). Все реакции пентозофосфатного пути
проходят в цитозоле клетки. Реакции неокислительного этапа пентозофосфатного
пути являются обратимыми, поэтому становится возможным синтез гексоз из пентоз.
Некоторые метаболиты неокислительного пути являются также и метаболитами
гликолиза. Из этого следует, что оба процесса тесно связаны и в зависимости от
потребностей клетки возможны переключения с одного пути на другой. При
сбалансированной потребности в NADPH и пентозах в клетке происходит
окислительный путь синтеза пентоз. Если потребности в пентозах превышают
потребности в NADPH, то окислительный путь шунтируется за счет использования
метаболитов гликолиза: фруктозо-6-фосфат и глицероальдегидфосфат в реакциях
неокислительного пути превращаются в пентозы.
Коферментом дегидрогеназ является
NADP+, который восстанавливается в NADPH и используется клетками в
реакции восстановления и гидроксилирования. Кроме того,
пентозофосфатный путь (окислительный и неокислительный) поставляет клетке пентозофосфаты,
необходимые для синтеза нуклеиновых кислот и
коферментов (NAD, FAD, СоА). Все реакции пентозофосфатного пути
проходят в цитозоле клетки. Реакции неокислительного этапа пентозофосфатного
пути являются обратимыми, поэтому становится возможным синтез гексоз из пентоз.
Некоторые метаболиты неокислительного пути являются также и метаболитами
гликолиза. Из этого следует, что оба процесса тесно связаны и в зависимости от
потребностей клетки возможны переключения с одного пути на другой. При
сбалансированной потребности в NADPH и пентозах в клетке происходит
окислительный путь синтеза пентоз. Если потребности в пентозах превышают
потребности в NADPH, то окислительный путь шунтируется за счет использования
метаболитов гликолиза: фруктозо-6-фосфат и глицероальдегидфосфат в реакциях
неокислительного пути превращаются в пентозы. Если же NADPH необходим
в большей степени, чем пентозы, то возможны два варианта:
Если же NADPH необходим
в большей степени, чем пентозы, то возможны два варианта:
При высоком энергетическом статусе клетки излишки пентоз путем обратных реакций неокислительного пути превращаются в фруктозо-6-фосфат и глицероальдегидфосфат, из которых в процессе глюконеогенеза образуется глюкоза;
При низком энергетическом статусе клетки из пентоз также образуются глицероальдегидфосфат и фруктозо-6-фосфат, которые затем включаются в гликолиз
Регуляция метаболизма углеводов (некоторые аспекты)
Регуляция метаболизма глюкозы в печени, связанная с
ритмом питания. Направление метаболизма глюкозы меняется при смене
периода пищеварения на постабсорбтивное состояние. При пищеварении глюкоза
задерживается в печени и депонируется в виде гликогена. Кроме того, глюкоза
используется для синтеза жиров. Причем, исходные субстраты для синтеза жира — a
-глицерофосфат и ацетил-СоА образуются из глюкозы в процессе гликолиза. Следовательно,
гликолиз в печени имеет особое значение. В постабсорбтивном периоде направление
процессов меняется на распад гликогена и глюконеогенез. Координация процессов
осуществляется путем аллостерической регуляции и ковалентной модификацией
ферментов (фосфорилирование и дефосфорилирование). Регуляторные
влияния направлены на реакции субстратных циклов. Активность ферментов
субстратного цикла фруктозо-6-фосфат « фруктозо-1,6-бисфосфат (цикл II) зависит
от концентрации фруктозо-2,6-бисфосфата, образующегося из фруктозо-6-фосфата в
дополнительной реакции. Синтез и распад этого регуляторного метаболита
происходят в реакциях, составляющих еще один субстратный цикл, оба направления
которого катализируются одним — бифункциональным ферментом (БИФ).
Киназная или фосфатазная активность
бифункционального фермента зависит от фосфорилированного или
дефосфорилированного состояния этого фермента.
Следовательно,
гликолиз в печени имеет особое значение. В постабсорбтивном периоде направление
процессов меняется на распад гликогена и глюконеогенез. Координация процессов
осуществляется путем аллостерической регуляции и ковалентной модификацией
ферментов (фосфорилирование и дефосфорилирование). Регуляторные
влияния направлены на реакции субстратных циклов. Активность ферментов
субстратного цикла фруктозо-6-фосфат « фруктозо-1,6-бисфосфат (цикл II) зависит
от концентрации фруктозо-2,6-бисфосфата, образующегося из фруктозо-6-фосфата в
дополнительной реакции. Синтез и распад этого регуляторного метаболита
происходят в реакциях, составляющих еще один субстратный цикл, оба направления
которого катализируются одним — бифункциональным ферментом (БИФ).
Киназная или фосфатазная активность
бифункционального фермента зависит от фосфорилированного или
дефосфорилированного состояния этого фермента. Концентрация
фруктозо-2,6-бисфосфата при пищеварении повышается, так как БИФ в этом случае
дефосфорилирован и проявляет киназную активность. Фруктозо-2,6-бисфосфат
является аллостерическим активатором гликолитического фермента и ингибитором
фермента глюконеогенеза. Следовательно, при пищеварении ускоряется
гликолитическое направление цикла и тормозится направление глюконеогенеза.
Фруктозо-1,6-дифосфат служит аллостерическим активатором пируваткиназы
(гликолитический фермент III цикла). В период пищеварения фруктозо-2,6-дифосфат
активирует фосфофруктокиназу и концентрация фруктозо-1,6-дифосфата
увеличивается, что приводит к активации пируваткиназы. Так достигается
согласованность в регуляции двух субстратных циклов. Регуляция I субстратного
цикла достигается следующим образом: при пищеварении концентрация глюкозы
повышается до 10-20 мкмоль/л. Активность глюкокиназы в этих условиях
максимальна, и глюкозо-6-фосфат направляется на синтез гликогена и жиров.
Концентрация
фруктозо-2,6-бисфосфата при пищеварении повышается, так как БИФ в этом случае
дефосфорилирован и проявляет киназную активность. Фруктозо-2,6-бисфосфат
является аллостерическим активатором гликолитического фермента и ингибитором
фермента глюконеогенеза. Следовательно, при пищеварении ускоряется
гликолитическое направление цикла и тормозится направление глюконеогенеза.
Фруктозо-1,6-дифосфат служит аллостерическим активатором пируваткиназы
(гликолитический фермент III цикла). В период пищеварения фруктозо-2,6-дифосфат
активирует фосфофруктокиназу и концентрация фруктозо-1,6-дифосфата
увеличивается, что приводит к активации пируваткиназы. Так достигается
согласованность в регуляции двух субстратных циклов. Регуляция I субстратного
цикла достигается следующим образом: при пищеварении концентрация глюкозы
повышается до 10-20 мкмоль/л. Активность глюкокиназы в этих условиях
максимальна, и глюкозо-6-фосфат направляется на синтез гликогена и жиров. Кроме
того, глюкоза при такой концентрации участвует в активации гликогенсинтазы.
Кроме
того, глюкоза при такой концентрации участвует в активации гликогенсинтазы.
Активность пируватдегидрогеназного комплекса в период пищеварения
также повышается, потому что он в этих условиях дефосфорилирован. В результате
ускоряется образование Ацетил-СоА и использование его для синтеза жирных
кислот. Переход ферментов из дефосфорилированного состояния в фосфорилированное
находится под контролем гормонов, в случае регуляции метаболизма глюкозы в
печени основными являются глюкагон и инсулин. Регуляция метаболизма глюкозы в
мышцах, связанная с режимом мышечной работы. При переходе от состояния покоя к
мышечной работе возрастает потребность клеток в энергии, которая восполняется
за счет ускорения процессов распада гликогена и гликолиза. Координация
интенсивности гликолиза обеспечивается аллостерической регуляцией ферментов
необратимых стадий энергетическим статусом клетки. Так, ингибитором
фосфофруктокиназы служит ATP, если в ходе гликолиза синтез ATP превышает
потребности клетки. В основе регуляции обмена гликогена лежит изменение
активности ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы. Регуляция
активности этих ферментов осуществляется путем фосфорилирования –
дефосфорилирования. Соотношение процессов синтеза гликогена, распада гликогена
и гликолиза в мышцах контролируют инсулин и адреналин.
В основе регуляции обмена гликогена лежит изменение
активности ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы. Регуляция
активности этих ферментов осуществляется путем фосфорилирования –
дефосфорилирования. Соотношение процессов синтеза гликогена, распада гликогена
и гликолиза в мышцах контролируют инсулин и адреналин.
Пути регуляции метаболизма углеводов крайне
разнообразны. На любых уровнях организации живого организма обмен углеводов регулируется
факторами, влияющими на активность
ферментов, участвующих в
реакциях углеводного обмена. К этим факторам относятся концентрация субстратов, содержание
продуктов (метаболитов) отдельных реакций, кислородный режим, температура, проницаемость
биологических мембран, концентрация коферментов, необходимых
для отдельных реакций, и т.д. (см. главу 4). В данной главе было
показано влияние перечисленных факторов на активность ферментных
систем углеводного обмена. И тем не менее некоторые
аспекты регуляции метаболизма
углеводов напомним.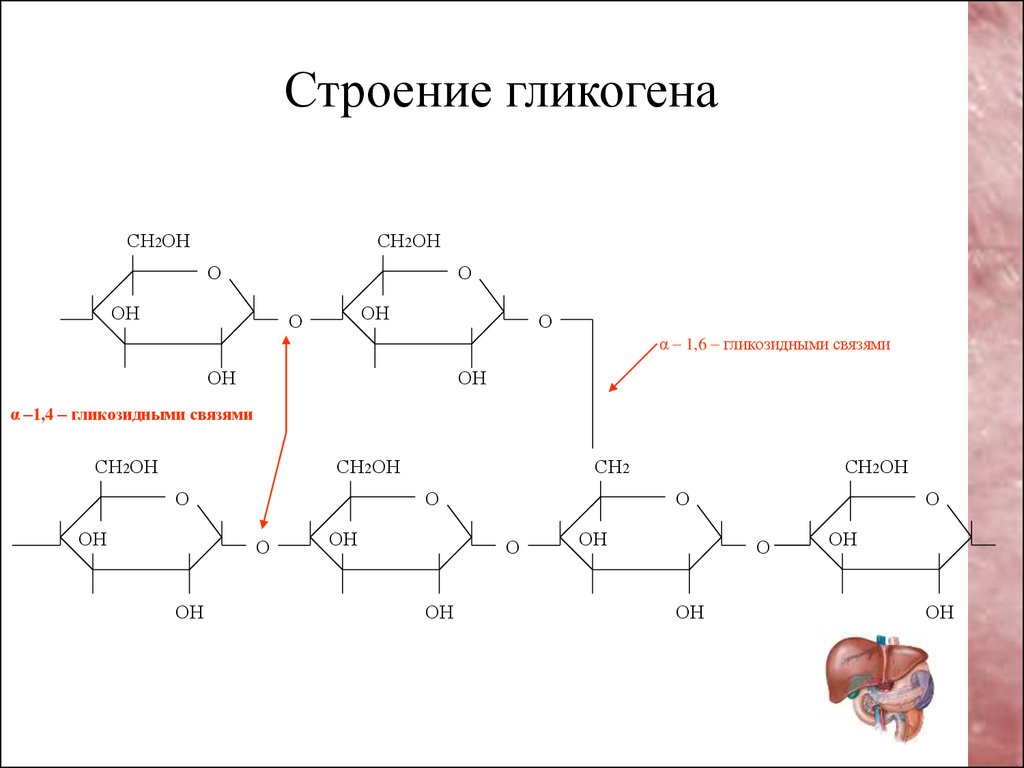 Гликолиз – это
совокупность реакций превращения глюкозы в пируват. У
аэробных организмов
гликолиз служит как бы
прелюдией к циклу
трикарбоновых кислот (циклу Кребса). Десять реакций гликолиза протекают в
цитозоле. Гликолитический путь играет двоякую роль: приводит к генерированию АТФ в результате распада глюкозы, и он же
поставляет строительные блоки для синтеза клеточных компонентов. Реакции
гликолитического пути в физиологических условиях легкообратимы, кроме реакций,
катализируемых гексокиназой,
фосфофруктокиназой и пируват-киназой. Фосфофруктокиназа – наиболее важный
регуляторный элемент (фермент) в процессе гликолиза, ингибируется
высокими концентрациями
АТФ и цитрата и активируется АМФ. Цикл трикарбоновых кислот
(цикл Кребса) представляет собой конечный общий путь для окисления «топливных» молекул. Большинство
«топливных» молекул
вступает в цикл в виде ацетил-КоА. Окислительное декарбоксилирование
пирувата, приводящее к образованию ацетил-КоА, является связующим звеном между гликолизом и циклом трикарбоновых кислот.
Гликолиз – это
совокупность реакций превращения глюкозы в пируват. У
аэробных организмов
гликолиз служит как бы
прелюдией к циклу
трикарбоновых кислот (циклу Кребса). Десять реакций гликолиза протекают в
цитозоле. Гликолитический путь играет двоякую роль: приводит к генерированию АТФ в результате распада глюкозы, и он же
поставляет строительные блоки для синтеза клеточных компонентов. Реакции
гликолитического пути в физиологических условиях легкообратимы, кроме реакций,
катализируемых гексокиназой,
фосфофруктокиназой и пируват-киназой. Фосфофруктокиназа – наиболее важный
регуляторный элемент (фермент) в процессе гликолиза, ингибируется
высокими концентрациями
АТФ и цитрата и активируется АМФ. Цикл трикарбоновых кислот
(цикл Кребса) представляет собой конечный общий путь для окисления «топливных» молекул. Большинство
«топливных» молекул
вступает в цикл в виде ацетил-КоА. Окислительное декарбоксилирование
пирувата, приводящее к образованию ацетил-КоА, является связующим звеном между гликолизом и циклом трикарбоновых кислот. Заметим, что последний служит также источником строительных блоков для
процессов биосинтеза.
Все реакции цикла протекают в митохондриях. Скорость цикла трикарбоновых кислот
зависит от потребности в АТФ.
Высокий энергетический заряд клетки понижает активность цитратсинтазы,
изоцитратдегидрогеназы и α-кетоглутаратдегидрогеназы.
Еще один важный регуляторный момент – необратимое образование ацетил-КоА из
пи-рувата. В результате пентозофосфатного пути происходит генерирование НАДФН и
рибозо-5-фосфата в цитозоле. НАДФН участвует в
восстановительных биосинтезах,
а рибозо-5-фосфат используется в синтезах РНК, ДНК и нуклеотидных коферментов. Взаимодействие гликолитического и
пентозофосфатного путей обеспечивает возможность постоянного приспособления концентраций НАДФН, АТФ и строительных блоков,
например рибозо-5-фосфата и пирувата, для удовлетворения потребностей клеток. Наконец, глюконеогенез и гликолиз регулируются
реципрокно, так что, если активность
одного из путей относительно понижается, то активность другого пути
повышается.
Заметим, что последний служит также источником строительных блоков для
процессов биосинтеза.
Все реакции цикла протекают в митохондриях. Скорость цикла трикарбоновых кислот
зависит от потребности в АТФ.
Высокий энергетический заряд клетки понижает активность цитратсинтазы,
изоцитратдегидрогеназы и α-кетоглутаратдегидрогеназы.
Еще один важный регуляторный момент – необратимое образование ацетил-КоА из
пи-рувата. В результате пентозофосфатного пути происходит генерирование НАДФН и
рибозо-5-фосфата в цитозоле. НАДФН участвует в
восстановительных биосинтезах,
а рибозо-5-фосфат используется в синтезах РНК, ДНК и нуклеотидных коферментов. Взаимодействие гликолитического и
пентозофосфатного путей обеспечивает возможность постоянного приспособления концентраций НАДФН, АТФ и строительных блоков,
например рибозо-5-фосфата и пирувата, для удовлетворения потребностей клеток. Наконец, глюконеогенез и гликолиз регулируются
реципрокно, так что, если активность
одного из путей относительно понижается, то активность другого пути
повышается. У человека и животных на всех стадиях синтеза и распада углеводов регуляция
углеводного обмена осуществляется при участии ЦНС и гормонов. Например,
установлено, что концентрация
глюкозы в крови ниже
3,3–3,4 ммоль/л (60–70 мг/100 мл) приводит к рефлекторному возбуждению высших
метаболических центров, расположенных в гипоталамусе. В регуляции углеводного
обмена особая роль принадлежит высшему отделу ЦНС – коре большого мозга. Наряду
с ЦНС важное влияние на содержание глюкозы оказывают
гормональные факторы, т.е. регуляции уровня глюкозы в крови осуществляется
ЦНС через ряд эндокринных желез
(см. главу 8).
У человека и животных на всех стадиях синтеза и распада углеводов регуляция
углеводного обмена осуществляется при участии ЦНС и гормонов. Например,
установлено, что концентрация
глюкозы в крови ниже
3,3–3,4 ммоль/л (60–70 мг/100 мл) приводит к рефлекторному возбуждению высших
метаболических центров, расположенных в гипоталамусе. В регуляции углеводного
обмена особая роль принадлежит высшему отделу ЦНС – коре большого мозга. Наряду
с ЦНС важное влияние на содержание глюкозы оказывают
гормональные факторы, т.е. регуляции уровня глюкозы в крови осуществляется
ЦНС через ряд эндокринных желез
(см. главу 8).
Цитохимические исследования содержания гликогена в лимфоцитах в лимфоцитарных пролиферациях | Кровь
Пропустить пункт назначенияСтатья| 1 августа 1958 г.
В. Дж. МИТУС, Л. Й. БЕРГНА, И. Б. МЕДНИКОФФ, В. ДАМЕШЕК
ДАМЕШЕККровь (1958) 13 (8): 748–756.
https://doi.org/10.1182/blood.V13.8.748.748
- Разделенный экран
- Делиться
- Фейсбук
- Твиттер
- Электронная почта
- Инструменты
Запросить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
- Открой PDF для в другом окне
Цитирование
В. Дж. МИТУС, Л. Дж. БЕРГНА, И. Б. МЕДНИКОФФ, В. ДАМЕШЕК; Цитохимические исследования содержания гликогена в лимфоцитах в лимфоцитарных пролиферациях. Кровь 1958; 13 (8): 748–756. doi: https://doi.org/10.1182/blood.V13.8.748.748
Дж. МИТУС, Л. Дж. БЕРГНА, И. Б. МЕДНИКОФФ, В. ДАМЕШЕК; Цитохимические исследования содержания гликогена в лимфоцитах в лимфоцитарных пролиферациях. Кровь 1958; 13 (8): 748–756. doi: https://doi.org/10.1182/blood.V13.8.748.748
Скачать файл цитирования:
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- Конечная примечание
- РефВоркс
- Бибтекс
Полуколичественным цитохимическим методом изучено содержание гликогена в лимфоцитах 44 случаев различных лимфоцитарных пролифераций. В лимфоцитах хронического лимфоцитарного лейкоза и лимфосаркомы (за одним исключением) значительно повышено содержание гликогена. При остром лимфолейкозе и в одном случае лимфосаркомы содержание гликогена было низким. Наличие или отсутствие клеток лимфосаркомы в периферической крови при лимфосаркоме не влияло на общий балл. Различие между гликоген-положительной «клеткой лимфосаркомы» и гликоген-отрицательным лимфобластом, по-видимому, связано с большей зрелостью цитоплазмы клетки лимфосаркомы (что является результатом асинхронизма ядерно-цитоплазматического созревания) и незрелостью цитоплазмы лимфобласта.
Наличие или отсутствие клеток лимфосаркомы в периферической крови при лимфосаркоме не влияло на общий балл. Различие между гликоген-положительной «клеткой лимфосаркомы» и гликоген-отрицательным лимфобластом, по-видимому, связано с большей зрелостью цитоплазмы клетки лимфосаркомы (что является результатом асинхронизма ядерно-цитоплазматического созревания) и незрелостью цитоплазмы лимфобласта.
Три случая доброкачественного лимфоцитоза с атипичными лимфоцитами в периферической крови дали повышенные значения. Один случай с атипичными лимфоцитами имел нормальные значения, и два случая лимфоцитоза с морфологически нормальными лимфоцитами также имели нормальные значения. Сделан вывод, что увеличение гликогена в лимфоцитарных клетках обусловлено общей пролиферацией лимфоцитов и не обязательно связано со злокачественным новообразованием как таковым.
Этот контент доступен только в формате PDF.Войдите через свое учреждение
Лабораторные исследования, визуализирующие исследования, другие тесты
 webmd.com»> von Gierke E. Гепато-нефромегалия гликогенная (Glykogenspeicherkrankheit der Leber und Nieren). Бейтр Путь Анат . 1929. 82:497-513.
webmd.com»> von Gierke E. Гепато-нефромегалия гликогенная (Glykogenspeicherkrankheit der Leber und Nieren). Бейтр Путь Анат . 1929. 82:497-513.Кори GT, Кори CF. Глюкозо-6-фосфатаза печени при болезни накопления гликогена. J Биол Хим . 1952 г., декабрь 199 (2): 661-7. [Ссылка QxMD MEDLINE].
Нарисава К., Игараси Ю., Отомо Х., Тада К. Новый вариант болезни накопления гликогена типа I, вероятно, связанный с дефектом транспортной системы глюкозо-6-фосфата. Biochem Biophys Res Commun . 1978 г., 29 августа. 83(4):1360-4. [Ссылка QxMD MEDLINE].
Джорджиева М., Раффин М., Дюшам А. и др. Прогрессирующее развитие кист почек при болезни накопления гликогена I типа. Hum Mol Genet . 2016 1 сентября. 25 (17): 3784-97. [Ссылка QxMD MEDLINE].
 webmd.com»> Дин Дж., Хуан Ю., Ян Х., Чжан Ц., Хоу С., Лю С. и др. [Анализ клинических особенностей 6 пациентов с болезнью накопления гликогена инфантильного типа II типа]. Чжунхуа Эр Кэ За Чжи . 2015 июнь 53 (6): 436-41. [Ссылка QxMD MEDLINE].
webmd.com»> Дин Дж., Хуан Ю., Ян Х., Чжан Ц., Хоу С., Лю С. и др. [Анализ клинических особенностей 6 пациентов с болезнью накопления гликогена инфантильного типа II типа]. Чжунхуа Эр Кэ За Чжи . 2015 июнь 53 (6): 436-41. [Ссылка QxMD MEDLINE].Dagli A, Sentner CP, Weinstein DA, Pagon RA, Adam MP, Ardinger HH, et al. Болезнь накопления гликогена III типа. 9 марта 2010 г. 1993-2017 гг. [Ссылка QxMD MEDLINE]. [Полный текст].
Сентнер С.П., Хугевен И.Дж., Вайнштейн Д.А., Сантер Р., Мерфи Э., Маккирнан П.Дж. и соавт. Болезнь накопления гликогена III типа: диагностика, генотип, лечение, клиническое течение и исход. J Наследовать Metab Dis . 2016 Сентябрь 39 (5): 697-704. [Ссылка QxMD MEDLINE].
Декостр В., Лафоре П., Надай-Паклеза А., Де Антонио М., Левегл С., Оливье Г. и др. Поперечное ретроспективное исследование мышечной функции у пациентов с болезнью накопления гликогена III типа.
 Нервно-мышечное расстройство . 2016 Сентябрь 26 (9): 584-92. [Ссылка QxMD MEDLINE].
Нервно-мышечное расстройство . 2016 Сентябрь 26 (9): 584-92. [Ссылка QxMD MEDLINE].Малфатти Э., Барнериас С., Хедберг-Олдфорс С. и др. Новая нервно-мышечная форма болезни накопления гликогена IV типа с артрогрипозом, скованностью позвоночника и редкими полиглюкозановыми тельцами в мышцах. Нервно-мышечное расстройство . 2016 26 октября (10): 681-7. [Ссылка QxMD MEDLINE].
McArdle B. Миопатия вследствие дефекта распада мышечного гликогена. Клиническая наука . 1951. 10:13-33.
Ногалес-Гадеа Г., Годфри Р., Санталла А., Колл-Канти Дж., Пинтос-Морелл Г., Пинос Т. и др. Гены и непереносимость физических упражнений: выводы из болезни МакАрдла. Физиол Геномикс . 2016 февраль 48 (2): 93-100. [Ссылка QxMD MEDLINE].
 com»> Grünert SC, Hannibal L, Spiekerkoetter U. Фенотипический и генетический спектр болезни накопления гликогена типа VI. Гены (Базель) . 3 августа 2021 г., 12 (8): [Ссылка на MEDLINE QxMD]. [Полный текст].
com»> Grünert SC, Hannibal L, Spiekerkoetter U. Фенотипический и генетический спектр болезни накопления гликогена типа VI. Гены (Базель) . 3 августа 2021 г., 12 (8): [Ссылка на MEDLINE QxMD]. [Полный текст].Ее HG. Дефицит альфа-глюкозидазы при генерализованной болезни накопления гликогена (болезнь Помпе). Биохим J . 1963 янв. 86:11-6. [Ссылка QxMD MEDLINE].
Таури С., Окуно Г., Икура Ю., Танака Т., Суда М., Нишикава М. Дефицит фосфофруктокиназы в скелетных мышцах. Новый тип гликогеноза. Biochem Biophys Res Commun . 1965 г., 3 мая. 19: 517-23. [Ссылка QxMD MEDLINE].
Derks TGJ, van Rijn M. Липиды при заболеваниях, связанных с накоплением гликогена в печени: патофизиология, мониторинг диетического питания и будущие направления. Журнал наследственных метаболических заболеваний .
 30 января 2015 г. 38(3):537-543. [Полный текст].
30 января 2015 г. 38(3):537-543. [Полный текст].Фуллер М., Даплок С., Тернер С. и др. Масс-спектрометрический количественный анализ гликогена для оценки накопления первичного субстрата у мышей Pompe. Анальная биохимия . 2012 15 февраля. 421 (2): 759-63. [Ссылка QxMD MEDLINE].
Каплер Э.Дж., Бергер К.И., Лешнер Р.Т. и др. КОНСЕНСУСНЫЕ РЕКОМЕНДАЦИИ ПО ЛЕЧЕНИЮ БОЛЕЗНИ ПОМПЕ С ПОСЛЕДНИМ НАЧАЛОМ. 2012;45(3):319-333. doi: 10.1002 / mus.22329. Мышцы и нервы . 2012. 45(3):319-333. [Ссылка QxMD MEDLINE].
Озен Х. Болезни накопления гликогена: новые перспективы. Всемирный журнал гастроэнтерологии . 14 мая 2007 г. 13:2541-2553. [Полный текст].
Магулас, PL, Эль-Хаттаб, AW. Болезнь накопления гликогена IV типа. Доступно по адресу https://www.
 ncbi.nlm.nih.gov/books/NBK115333/. 2013; Доступ: 15 мая 2017 г.
ncbi.nlm.nih.gov/books/NBK115333/. 2013; Доступ: 15 мая 2017 г.Мартин М.А., Лусия А., Аренас Дж., Андреу А.Л., Пагон Р.А., Адам М.П. и др. Болезнь накопления гликогена типа V. 19 апреля 2006 г. [Ссылка QxMD MEDLINE]. [Полный текст].
Кришнамурти Н., Сантош В., Яша Т.К., Махадеван А., Шанкар С.К., Джетвани Д. и др. Болезнь накопления гликогена V типа (болезнь Мак-Ардла): отчет о трех случаях. Нейрол Индия . 2011 ноябрь-декабрь. 59(6):884-6. [Ссылка QxMD MEDLINE].
Musumeci O, Bruno C, Mongini T, Rodolico C, Aguennouz M, Barca E, et al. Клинические особенности и новые молекулярные данные при дефиците мышечной фосфофруктокиназы (GSD тип VII). Нервно-мышечное расстройство . 2011 29 ноября. [Ссылка на MEDLINE QxMD].
Эллингвуд С.
 С., Ченг А. Биохимические и клинические аспекты болезней накопления гликогена. J Эндокринол . 2018 сен. 238 (3): R131-R141. [Ссылка QxMD MEDLINE]. [Полный текст].
С., Ченг А. Биохимические и клинические аспекты болезней накопления гликогена. J Эндокринол . 2018 сен. 238 (3): R131-R141. [Ссылка QxMD MEDLINE]. [Полный текст].Бейзаи З., Герамизаде Б. Молекулярная диагностика болезни накопления гликогена I типа: обзор. ЭКСЛИ Дж . 2019. 18:30-46. [Ссылка QxMD MEDLINE].
Матерн Д., Зейдевиц Х.Х., Бали Д., Ланг С., Чен Ю.Т. Болезнь накопления гликогена I типа: диагностика и корреляция фенотип/генотип. Евро J Педиатр . 2002 г., октябрь 161, Приложение 1: S10-9. [Ссылка QxMD MEDLINE].
Мелис Д., Фульсери Р., Паренти Г. и др. Корреляция генотипа/фенотипа при болезни накопления гликогена типа 1b: многоцентровое исследование и обзор литературы. Евро J Педиатр . 2005 авг. 164(8):501-8. [Ссылка QxMD MEDLINE].
 webmd.com»> Kuijpers TW, Maianski NA, Tool AT, et al. Апоптотические нейтрофилы в кровотоке у пациентов с болезнью накопления гликогена типа 1b (GSD1b). Кровь . 2003 15 июня. 101 (12): 5021-4. [Ссылка QxMD MEDLINE].
webmd.com»> Kuijpers TW, Maianski NA, Tool AT, et al. Апоптотические нейтрофилы в кровотоке у пациентов с болезнью накопления гликогена типа 1b (GSD1b). Кровь . 2003 15 июня. 101 (12): 5021-4. [Ссылка QxMD MEDLINE].Cheung YY, Kim SY, Yiu WH, et al. Нарушенная активность нейтрофилов и повышенная восприимчивость к бактериальным инфекциям у мышей с дефицитом глюкозо-6-фосфатазы-бета. Дж Клин Инвест . 2007 март 117(3):784-93. [Ссылка QxMD MEDLINE].
Janecke AR, Bosshard NU, Mayatepek E, et al. Молекулярная диагностика болезни накопления гликогена 1с. Гум Жене . 1999 март 104(3):275-7. [Ссылка QxMD MEDLINE].
Шигето С., Катафути Т., Окада Ю., Накамура К., Эндо Ф., Окуяма Т. и др. Усовершенствованный анализ для дифференциальной диагностики между болезнью Помпе и псевдодефицитом кислой α-глюкозидазы на высохших пятнах крови.
 Мол Генет Метаб . 2011 май. 103 (1):12-7. [Ссылка QxMD MEDLINE].
Мол Генет Метаб . 2011 май. 103 (1):12-7. [Ссылка QxMD MEDLINE].Ногалес-Гадеа Г., Брюлл А., Санталла А., Андреу А.Л., Аренас Дж., Мартин М.А. и др. Болезнь Макардла: обновление зарегистрированных мутаций и полиморфизмов в гене PYGM. Хум Мутат . 2015 36 июля (7): 669-78. [Ссылка QxMD MEDLINE].
Димаур С., Андреу А.Л., Бруно С., Хаджигеоргиу Г.М. Дефицит миофосфорилазы (гликогеноз V типа, болезнь Мак-Ардла). Карр Мол Мед . 2002 март 2 (2): 189-96. [Ссылка QxMD MEDLINE].
Ховард Т.Д., Акоц Г., Боуден Д.В. Физическое и генетическое картирование гена мышечной фосфофруктокиназы (PFKM): переназначение на хромосому 12q человека. Геномика . 1996 г., 15 мая. 34(1):122-7. [Ссылка QxMD MEDLINE].
Джейкоби Дж.
 Т., Бенто Дос Сантос Б., Налин Т., Колонетти К., Фаррет Рефоско Л., Ф.М. де Соуза С. и др. Минеральная плотность костей у пациентов с заболеваниями печени, связанными с накоплением гликогена. Питательные вещества . 2021 авг. 27. 13 (9): 2987. [Ссылка QxMD MEDLINE]. [Полный текст].
Т., Бенто Дос Сантос Б., Налин Т., Колонетти К., Фаррет Рефоско Л., Ф.М. де Соуза С. и др. Минеральная плотность костей у пациентов с заболеваниями печени, связанными с накоплением гликогена. Питательные вещества . 2021 авг. 27. 13 (9): 2987. [Ссылка QxMD MEDLINE]. [Полный текст].Бали Д.С., Чен Ю.Т., Остин С., Гольдштейн Дж.Л., Пагон Р.А., Адам М.П. и др. Болезнь накопления гликогена типа I. GeneReviews . Август 2016 г. [Ссылка QxMD MEDLINE]. [Полный текст].
Чен Ю.Т. Болезнь накопления гликогена I типа: поражение почек, патогенез и лечение. Педиатр Нефрол . 1991 5 января (1): 71–76. [Ссылка QxMD MEDLINE].
Манди Х.Р., Ли П.Дж. Гликогеноз I типа и сахарный диабет: общий механизм почечной дисфункции? Мед Гипотезы . 2002 г., июль 59 (1): 110-4. [Ссылка QxMD MEDLINE].

Пападимас Г.К., Спенгос К., Пападопулос С., Манта П. Болезнь накопления гликогена с поздним началом II типа: подводные камни в диагностике. Евро Нейрол . 2011 15 декабря. 67(2):65-68. [Ссылка QxMD MEDLINE].
Виссер Г., Рейк Д.П., Кокке Ф.Т., Никкельс П.Г., Зауэр П.Дж., Смит Г.П. Функция кишечника при болезни накопления гликогена типа I. J Inherit Metab Dis . 2002 г. 25 августа (4): 261-7. [Ссылка QxMD MEDLINE].
Лесли Н., Бейли Л. Болезнь Помпе. Доступно на https://www.ncbi.nlm.nih.gov/books/NBK1261/. 31 августа 2007 г. [Обновлено 11 мая 2017 г.]; Доступ: 15 мая 2017 г.
Рабочая группа по диагностике болезни Помпе., Winchester B, Bali D, Bodamer OA, et al. Методы быстрой и надежной лабораторной диагностики болезни Помпе: отчет с международного консенсусного совещания.
Мол Генет Метаб . 2008 март 93 (3): 275-81. [Ссылка QxMD MEDLINE].Кишнани П.С., Штайнер Р.Д., Бали Д., Бергер К., Бирн Б.Дж., Кейс Л.Е. и др. Руководство по диагностике и лечению болезни Помпе. Жене Мед . 2006 май. 8 (5): 267-88. [Ссылка QxMD MEDLINE].
Chien YH, Lee NC, Thurberg BL, Chiang SC, Zhang XK, Keutzer J, et al. Болезнь Помпе у младенцев: улучшение прогноза путем скрининга новорожденных и раннего лечения. Педиатрия . 2009 г., декабрь 124 (6): e1116-25. [Ссылка QxMD MEDLINE].
Мехтлер Т.П., Стари С., Мец Т.Ф., Де Хесус В.Р., Гребер-Платцер С., Поллак А. и др. Неонатальный скрининг на лизосомные болезни накопления: осуществимость и заболеваемость по результатам общенационального исследования в Австрии. Ланцет . 2012 28 января. 379 (9813): 335-41. [Ссылка QxMD MEDLINE].
Chien YH, Chiang SC, Zhang XK, Keutzer J, Lee NC, Huang AC, et al. Раннее выявление болезни Помпе с помощью скрининга новорожденных возможно: результаты тайваньской программы скрининга. Педиатрия . 2008 г., июль 122 (1): e39-45. [Ссылка QxMD MEDLINE].
Мартин М.А., Лусия А., Аренас Дж., Андреу А.Л., Пагон Р.А., Адам М.П. и др. Болезнь накопления гликогена типа V. 1993. [Ссылка QxMD MEDLINE]. [Полный текст].
Кишнани П.С., Остин С.Л., Абденур Дж.Э., Арн П., Бали Д.С., Бони А. и др. Диагностика и лечение болезни накопления гликогена I типа: практическое руководство Американского колледжа медицинской генетики и геномики. Генет Мед . 2014 16 (11) ноября: e1. [Ссылка QxMD MEDLINE].
Деграсси И., Дехерагода М., Криген Д. и др.
Гистология печени у детей с нарушениями накопления гликогена VI и IX типа. Раскопать печень Dis . 2021 янв. 53 (1): 86-93. [Ссылка QxMD MEDLINE].Бали Д.С., Эль-Гарбави А., Остин С. и др. Болезнь накопления гликогена типа I. Адам М.П., Эверман Д.Б., Мирзаа Г.М. и др., ред. GeneReviews® [Интернет] . Сиэтл, Вашингтон: Вашингтонский университет; 2021. 1993-2022. [Полный текст].
Derks TGJ, Rodriguez-Buritica DF, Ahmad A, de Boer F, Couce ML, Grünert SC, et al. Болезнь накопления гликогена типа Ia: текущие варианты лечения, бремя и неудовлетворенные потребности. Питательные вещества . 2021 27 окт. 13 (11): 3828. [Ссылка QxMD MEDLINE]. [Полный текст].
Бхаттачарья К., Ортон Р.С., Ци С. и др. Новый крахмал для лечения болезней накопления гликогена. J Наследовать Metab Dis .
 2007 30 июня (3): 350-7. [Ссылка QxMD MEDLINE].
2007 30 июня (3): 350-7. [Ссылка QxMD MEDLINE].Росс К.М., Браун Л.М., Коррадо М.М., Ченгсупанимит Т., Карри Л.М., Ферреккья И.А. и др. Безопасность и эффективность хронической терапии кукурузным крахмалом с пролонгированным высвобождением при болезни накопления гликогена типа I. JIMD Rep . 2016. 26:85-90. [Ссылка QxMD MEDLINE].
Коберл Д.Д., Кишнани П.С., Чен Ю.Т. Болезнь накопления гликогена I и II типов: обновление лечения. J Наследовать Metab Dis . 2007 г., 30 апреля (2): 159–64. [Ссылка QxMD MEDLINE].
Кишнани П.С., Сан Б., Коберл Д.Д. Генная терапия болезней накопления гликогена. Хум Мол Жене . 2019 1 октября. 28 (R1): R31-R41. [Ссылка QxMD MEDLINE]. [Полный текст].
Чоу, Дж. Ю., Чо, Дж. Х., Ким, Г. Я. и другие.
 Молекулярная биология и генная терапия болезни накопления гликогена типа Ib. J Наследовать Metab Dis . 2018. 41:1007-1014. [Полный текст].
Молекулярная биология и генная терапия болезни накопления гликогена типа Ib. J Наследовать Metab Dis . 2018. 41:1007-1014. [Полный текст].Чжоу З., Остин Г.Л., Шаффер Р., Армстронг Д.Д., Джентри М.С. Ферментная терапия, опосредованная антителами, и ее применение при заболеваниях накопления гликогена. Тренды Мол Мед . 2019 25 декабря (12): 1094-1109. [Ссылка QxMD MEDLINE]. [Полный текст].
Чжан Л., Ли С., Арнаутова И. и др. Генная терапия с использованием нового варианта G6PC-S298C повышает долгосрочную эффективность лечения болезни накопления гликогена типа Ia. Biochem Biophys Res Commun . 2020 30 июня. 527 (3): 824-30. [Ссылка QxMD MEDLINE]. [Полный текст].
Арнаутова И., Чжан Л., Чен Х.Д., Мэнсфилд Б.К., Чоу Дж.Ю. Коррекция метаболических нарушений в мышиной модели болезни накопления гликогена типа Ia с помощью CRISPR/Cas9на основе редактирования генов.
 Мол Тер . 2021 7 апр. 29 (4): 1602-10. [Ссылка QxMD MEDLINE]. [Полный текст].
Мол Тер . 2021 7 апр. 29 (4): 1602-10. [Ссылка QxMD MEDLINE]. [Полный текст].Гарбаде С.Ф., Эдерер В., Бургард П. и др. Влияние болезни накопления гликогена I типа на повседневную жизнь взрослых: обзор. Orphanet J Rare Dis . 2021 3 сент. 16 (1): 371. [Ссылка QxMD MEDLINE]. [Полный текст].
Reuser AJ, Verheijen FW, Bali D, van Diggelen OP, Germain DP, Hwu WL и др. Использование высушенных образцов капель крови в диагностике лизосомных болезней накопления — современное состояние и перспективы. Мол Генет Метаб . 2011 сен-окт. 104 (1-2): 144-8. [Ссылка QxMD MEDLINE].
Кишнани П.С., Корзо Д., Лесли Н.Д., Грускин Д., Ван дер Плоэг А., Клэнси Дж.П. и др. Раннее начало лечения алглюкозидазой альфа продлевает долгосрочную выживаемость младенцев с болезнью Помпе. Pediatr Res .
 2009 сен. 66 (3): 329-35. [Ссылка QxMD MEDLINE].
2009 сен. 66 (3): 329-35. [Ссылка QxMD MEDLINE].Кишнани П.С., Гольденберг П.С., ДеАрми С.Л., Хеллер Дж., Бенджамин Д., Янг С. и др. Перекрестно-реактивный иммунологический статус материала влияет на результаты лечения детей с болезнью Помпе. Мол Генет Метаб . 2010 янв. 99 (1): 26-33. [Ссылка QxMD MEDLINE].
Мессингер Ю.Х., Мендельсон Н.Дж., Рхед В., Диммок Д., Хершковитц Э., Чемпион М. и др. Успешная индукция иммунной толерантности к заместительной ферментной терапии при CRIM-негативной детской болезни Помпе. Жене Мед . 2012 14 января (1): 135-42. [Ссылка QxMD MEDLINE].
Martin-Touaux E, Puech JP, Chateau D и др. Мышцы как предполагаемый продуцент кислой альфа-глюкозидазы для генной терапии гликогеноза II типа. Хум Мол Жене . 2002 1 июля. 11 (14): 1637-45. [Ссылка QxMD MEDLINE].

Lim JA, Choi SJ, Gao F, Kishnani PS, Sun B. Новый подход к генной терапии для GSD III с использованием вектора AAV, кодирующего бактериальный фермент, расщепляющий гликоген. Mol Ther Methods Clin Dev . 2020 11 сент. 18:240-9. [Ссылка QxMD MEDLINE]. [Полный текст].
Roe CR, Mochel F. Анаплеротическая диетотерапия при наследственных метаболических заболеваниях: терапевтический потенциал. J Наследовать Metab Dis . 2006 апрель-июнь. 29(2-3):332-40. [Ссылка QxMD MEDLINE].
Кинман Р.П., Касумов Т., Джоббинс К.А., Томас К.Р., Адамс Дж.Е., Бруненграбер Л.Н. Парентеральный и энтеральный метаболизм анаплеротического тригептаноина у нормальных крыс. Am J Physiol Endocrinol Metab . 2006 г., октябрь 291 (4): E860-6. [Ссылка QxMD MEDLINE].

