Обмен глюкозы и зачатие | Махмудов
Аннотация
Рождение полноценного жизнеспособного потомства находится в прямой зависимости от состояния материнского организма. Начиная с самых ранних периодов беременности многие факторы внешней и внутренней среды оказывают влияние на внутриутробное развитие плода. Успешное завершение беременности и рождение здорового потомства возможны при сбалансированности обмена веществ и удовлетворении всех потребностей материнского организма. В этом процессе большую роль играют углеводы, и в частности глюкоза, которая интенсивно используется внутриутробно развивающимся эмбрионом. Дефицит ее в организме матери может затормозить развитие и даже вызвать раннюю гибель потомства. Поэтому, учитывая важность глюкозы для развивающегося организма, в настоящем обзоре обсуждаются вопросы, связанные с ее обменом и регуляцией в процессе беременности.
Ключевые слова
глюкоза, инсулин, зачатие, эмбриогенез
Для цитирования:
Махмудов Э. С., Ходжиматов В.А. Обмен глюкозы и зачатие. Проблемы Эндокринологии. 1993;39(2):60-62.
С., Ходжиматов В.А. Обмен глюкозы и зачатие. Проблемы Эндокринологии. 1993;39(2):60-62.
For citation:
Makhmudov E.S., Khodzhimatov V.A. Glucose metabolism and conception. Problems of Endocrinology. 1993;39(2):60-62. (In Russ.)
Рождение полноценного жизнеспособного потомства находится в прямой зависимости от состояния материнского организма. Начиная с самых ранних периодов беременности многие факторы внешней и внутренней среды оказывают влияние на внутриутробное развитие плода. Успешное завершение беременности и рождение здорового потомства возможны при сбалансированности обмена веществ и удовлетворении всех потребностей материнского организма. В этом процессе большую роль играют углеводы, и в частности глюкоза, которая интенсивно используется внутриутробно развивающимся эмбрионом.
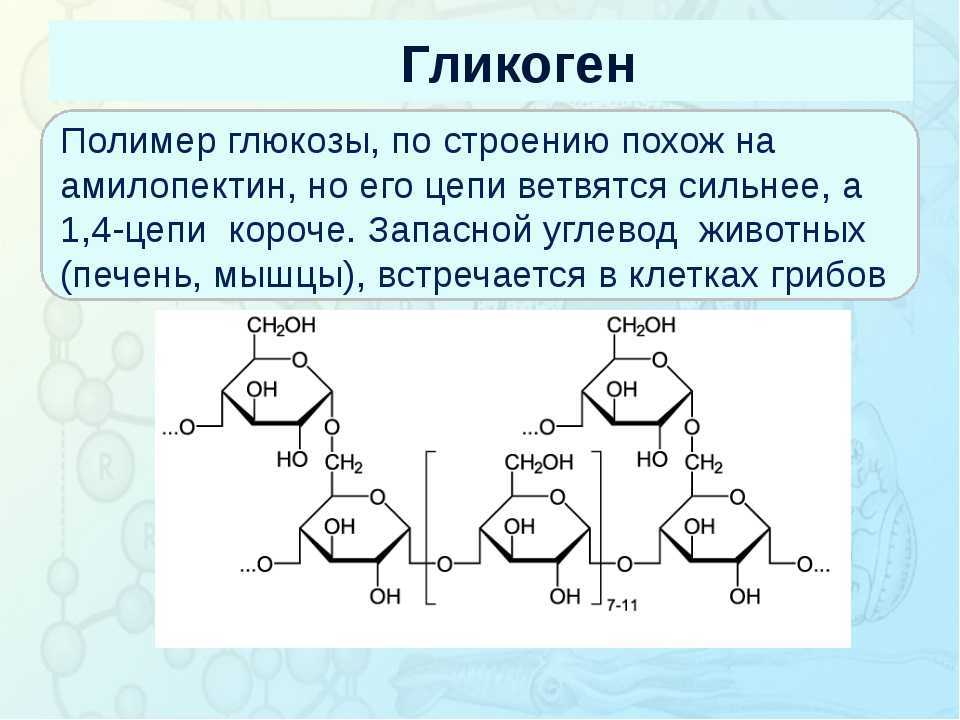
Глюкоза является одним из основных энергетических субстратов. В свободном виде она всегда присутствует в крови и тканевых жидкостях организма. Концентрация ее в периферической крови относится к константным показателям, у здорового человека в состоянии относительного покоя ее величина составляет 4,5—5,5 ммоль/л, а общее количество глюкозы в организме составляет приблизительно 20 г [11]. В процессе эволюции сформировалась сложная система регуляции глюкозы в крови [4].
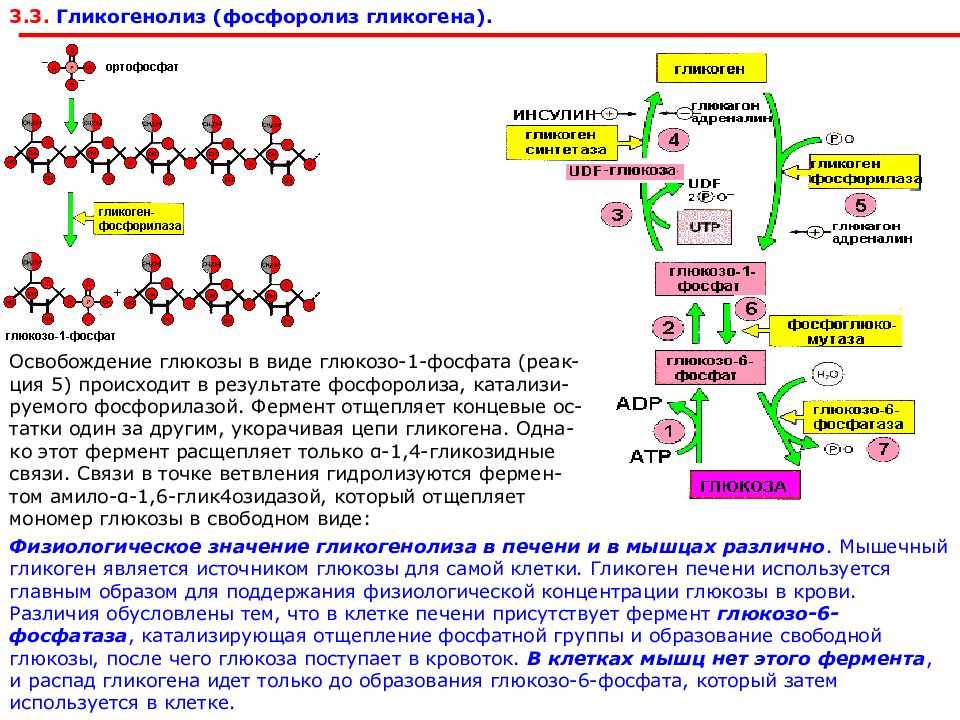
Глюкоза, поступающая в организм с пищей, может резервироваться в виде гликогена или использоваться для быстрого образования энергии, необходимой клеткам и тканям. Окисление глюкозы в тканях высших организмов сопровождается освобождением химической энергии, аккумулированной в форме АТФ.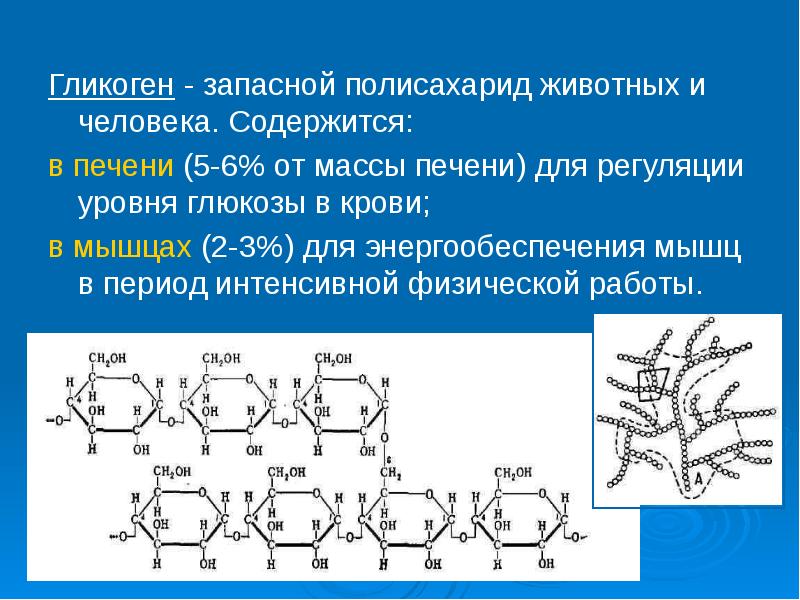 В процессе анаэробного распада глюкозы образуются 2—3 молекулы, а при аэробном, наиболее выгодном пути окисления, 38—39 молекул АТФ [2]. Использование глюкозы в организме зависит от скорости ее накопления в виде гликогена в мышцах и печени [8]. Гликоген печени по мере надобности быстро распадается с высвобождением глюкозы. Обмен углеводов может изменяться не более чем на 30 % при различных функциональных напряжениях организма человека и животных [6, 8].
В процессе анаэробного распада глюкозы образуются 2—3 молекулы, а при аэробном, наиболее выгодном пути окисления, 38—39 молекул АТФ [2]. Использование глюкозы в организме зависит от скорости ее накопления в виде гликогена в мышцах и печени [8]. Гликоген печени по мере надобности быстро распадается с высвобождением глюкозы. Обмен углеводов может изменяться не более чем на 30 % при различных функциональных напряжениях организма человека и животных [6, 8].
Это касается и беременности, в процессе которой наступает глубокая перестройка метаболических процессов в организме. Изменения, возникающие в организме матери, начиная с самых ранних сроков беременности, отражаются и на внутриутробно развивающемся потомстве, ибо между ними устанавливается тесная взаимосвязь [12, 13].
С наступлением беременности в крови животных нарастает уровень ряда гормонов (прогестерона, эстрадиола), усиливается кровоснабжение репродуктивных органов и существенно перестраивается обмен веществ в сторону накопления в организме матери жира [40].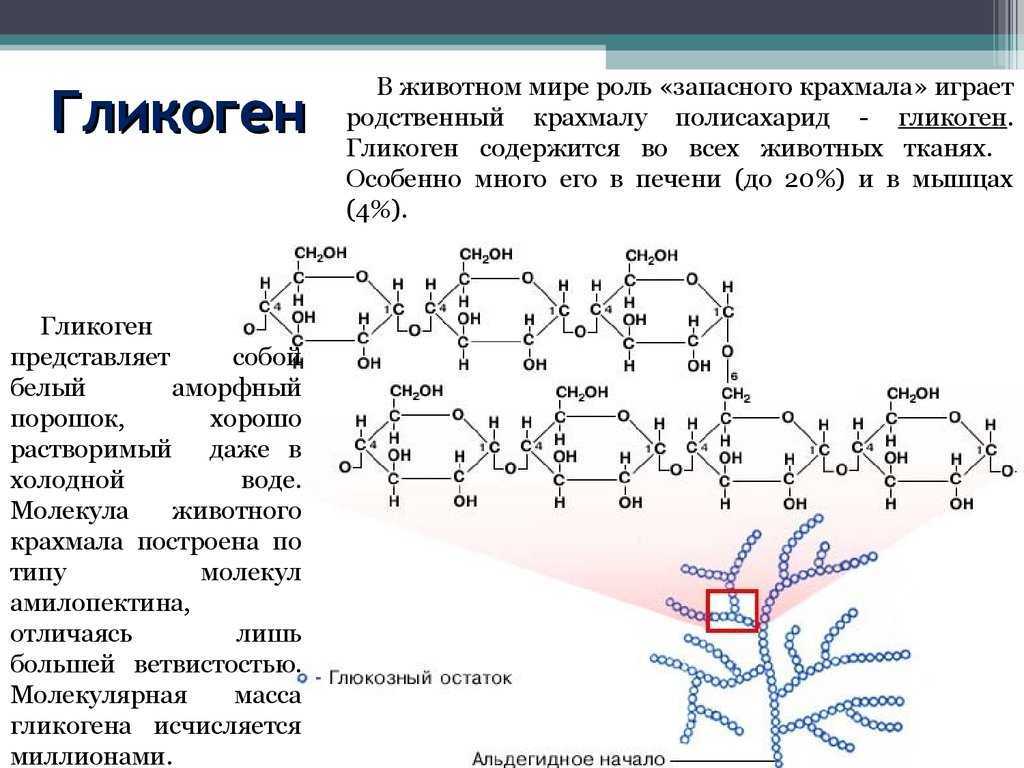 Одновременно активируется и обмен углеводов. Внутривенное введение беременным ‘’С-аланина, ‘4С-глицина или |4С-пирувата приводит к усиленному образованию нС-глюкозы в срезах печени [19]. О высокой интенсивности обмена глюкозы свидетельствуют и другие работы. Например, у крыс линии Rattus — Rattus во время беременности содержание гликогена в печени, скелетных мышцах и матке снижается, в то время как концентрация глюкозы в крови возрастает. Последняя используется для покрытия энергетических потребностей развивающихся эмбрионов [49]. Эксперименты, проведенные на беременных овцах, показали, что при нормогликемических состояниях скорость выработки глюкозы в 2 раза выше, чем у овец с гипогликемией. В матке крольчих утилизация глюкозы крови при беременности увеличивается [26].
Одновременно активируется и обмен углеводов. Внутривенное введение беременным ‘’С-аланина, ‘4С-глицина или |4С-пирувата приводит к усиленному образованию нС-глюкозы в срезах печени [19]. О высокой интенсивности обмена глюкозы свидетельствуют и другие работы. Например, у крыс линии Rattus — Rattus во время беременности содержание гликогена в печени, скелетных мышцах и матке снижается, в то время как концентрация глюкозы в крови возрастает. Последняя используется для покрытия энергетических потребностей развивающихся эмбрионов [49]. Эксперименты, проведенные на беременных овцах, показали, что при нормогликемических состояниях скорость выработки глюкозы в 2 раза выше, чем у овец с гипогликемией. В матке крольчих утилизация глюкозы крови при беременности увеличивается [26].
У человека и животных в период беременности содержание глюкозы в крови и гликогена в печени существенно снижается [26, 27, 54]. В одном из последних обзоров [41], посвященном метаболизму глюкозы, указывается, что у разных видов животных в поздние сроки беременности происходит снижение уровня глюкозы в крови при одновременном увеличении в ней .
Нами также проведено определение содержания глюкозы в крови крыс в разные периоды беременности в условиях относительного температурного комфорта и на фоне воздействия высокой окружающей температуры. Эксперименты показали, что, начиная с ранних сроков беременности, уровень глюкозы в крови снижается, но особенно резко после гипертермических воздействий на организм. В этих условиях скармливание глюкозы или ее внутривенное введение беременным животным улучшают углеводный обмен, повышают воспроизводительную функцию и жизнеспособность потомства [1, 9]. Эстрогены, введенные в ранние сроки беременности, стимулируют поступление глюкозы в полость матки и тем самым способствуют имплантации бластоцисты [45].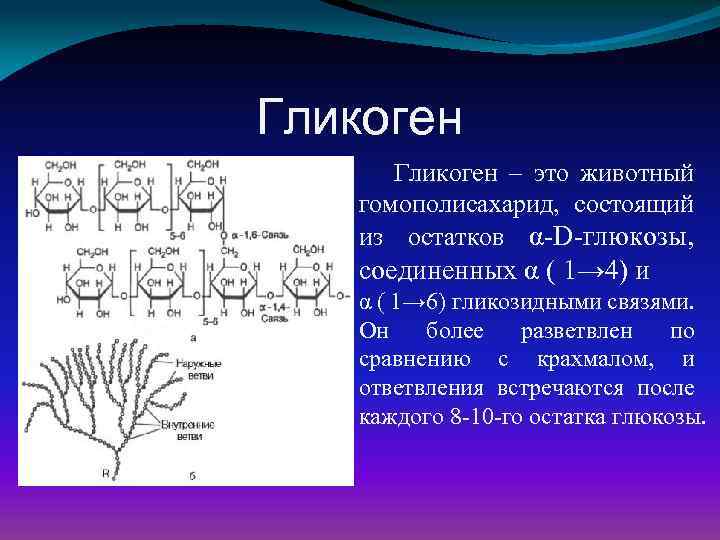
Имеются данные, что значительная часть глюкозы у эмбрионов свиньи до 8-й клеточной стадии развития обменивается по пентозофосфатному пути, а начиная с 8-й клеточной стадии до бластоцисты по гликолитическому пути [23]. В то же время на мышиных оплодотворенных яйцеклетках установлено, что глюкоза предпочтительно утилизируется в период имплантации и не используется до формирования бластоцисты. Однако на человеческих зародышах показано, что до стадии бластоцисты они используют глюкозу, другие эндогенные источники и возможно гликоген. Установлен факт влияния глюкозы на имплантацию у овец и крыс [1, 3].
В наших исследованиях также показано, что скармливание крысам в период имплантации глюкозы из расчета 1 г на 100 г массы сопровождается повышением рождаемости крыс на 20 % по сравнению с контрольными животными [10]. Аналогичную закономерность наблюдали и другие исследователи, вводившие в рацион беременных крыс нарастающие количества углеводов [37].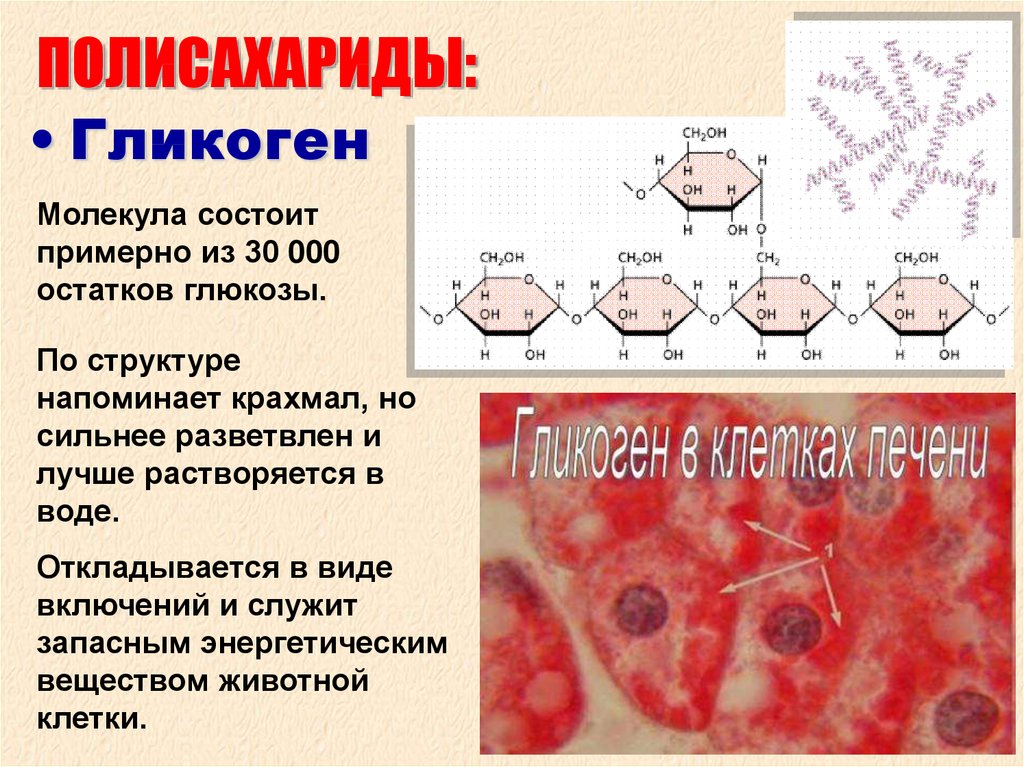
Интересно отметить, что на адгезивной стадии имплантации обнаруживаются изменения в продукции D-галактозы и N-ацетилгалактозы. Введение в просвет одного рога беременной матки мыши в предымплантационный период X-D-N-ацетилгалактозиламинидазы, р-галактозидазы или а-галактозидазы или лактальбумина предотвращает этот процесс у одних мышей и задерживает эмбриональное развитие у других. •
Следовательно, нарушение метаболизма О-галактозы мешает процессу имплантации и тормозит активность децидуальных клеток [21]. Также показано, что нарушение метаболизма углеводов, и в частности глюкозы, приводит к бесплодию. Из этого следует, что глюкоза в материнском организме является важным энергетическим субстратом, не только поддерживающим репродуктивную функцию, но и обеспечивающим метаболические потребности внутриутробно развивающегося эмбриона [41]. Использованию глюкозы способствует и ее свободное прохождение через плаценту путем облегченной диффузии [16]. Причем по мере внутриутробного развития потребность плода в энергетических субстратах возрастает, поэтому у беременных животных повышаются объем распределения глюкозы, ее ассимиляция и скорость распределения между легко- и труднообмени- ваемой фракциями [17]. О существенном значении глюкозы, содержащейся в крови матери, для роста и развития плода свидетельствуют эксперименты с перевязкой одного рога крысиной матки в период беременности. При этом уровень сахара в крови у матери не изменяется, в то время как у плода он достоверно снижается, а рост и развитие плода в перевязанном роге тормозятся [49]. Кроме глюкозы, зародыш или плод используют лактат, пируват и другие крмпоненты гликолиза и цикла лимонной кислоты. Кстати, в эмбриональной ткани активность ферментов анаэробного распада углеводов превалирует над активностью ферментов аэробного окисления. Этот путь превращения углеводов остается весьма активным во многих тканях новорожденного и ребенка 1-го месяца жизни [2].
Использованию глюкозы способствует и ее свободное прохождение через плаценту путем облегченной диффузии [16]. Причем по мере внутриутробного развития потребность плода в энергетических субстратах возрастает, поэтому у беременных животных повышаются объем распределения глюкозы, ее ассимиляция и скорость распределения между легко- и труднообмени- ваемой фракциями [17]. О существенном значении глюкозы, содержащейся в крови матери, для роста и развития плода свидетельствуют эксперименты с перевязкой одного рога крысиной матки в период беременности. При этом уровень сахара в крови у матери не изменяется, в то время как у плода он достоверно снижается, а рост и развитие плода в перевязанном роге тормозятся [49]. Кроме глюкозы, зародыш или плод используют лактат, пируват и другие крмпоненты гликолиза и цикла лимонной кислоты. Кстати, в эмбриональной ткани активность ферментов анаэробного распада углеводов превалирует над активностью ферментов аэробного окисления. Этот путь превращения углеводов остается весьма активным во многих тканях новорожденного и ребенка 1-го месяца жизни [2].
Исследования с меченой глюкозой показали, что значительная ее часть у плода окисляется в пентозофосфат — ном цикле. Глюкоза действует на специфический рецептор, активирующий аденилатциклазу, ведет к увеличению уровня аденозинмонофосфата, усиливает процессы трансформации энергии, аккумулированной в макроэргических связях. Образующаяся при этом энергия используется для нужд интенсивно растущего организма [15]. Начиная с 16-го дня внутриутробной жизни плод использует глюкозу и для синтеза гликогена [39]. Гепатоциты 18-дневных плодов способны интенсивно синтезировать гликоген, содержание которого при наличии следовых количеств инсулина повышается почти в 3 раза [42]. В этой связи представляется важным обсудить степень участия гормонов, в частности инсулина и глюкагона, в регуляции углеводного обмена в период беременности. Преобладающим числом исследований показано увеличение содержания инсулина в крови здоровых беременных женщин и животных [14, 27], что сопровождается снижением утилизации глюкозы инсулинчувствительными тканями матери, а ее использование развивающимся плодом постепенно увеличивается.
Уровень глюкозы в крови заметно нарастает сразу после еды и резко снижается у голодных животных. Нестабильность концентрации глюкозы в крови поддерживается плацентарным лактогеном и прогестероном, которые снижают чувствительность периферических тканей к инсулину и соответственно утилизацию в них углеводов [47, 51]. В адипоцитах беременных крыс снижаются поступление, превращение и окисление стимулируемой инсулином глюкозы [52]. В то же время инсулин и инсулиноподобные факторы роста повышают утилизацию глюкозы, образование лактата, СО? и накопление гликогена в тканях изолированной плаценты человека [49]. Считается, что гликогенсодержащие клетки выполняют защитную роль и дают возможность совместного существования клеток материнского и зародышевого происхождения внутри плаценты в течение беременности»]!)].
Помимо влияния на углеводный обмен, инсулин стимулирует образование инсулиноподобного ростового фактора 1 и оба гормона оказывают анаболическое действие на плод [30]. При этом циркулирующий в крови матери инсулин не проникает через плаценту, но от его секреции зависит содержание глюкозы в крови матери и поступление к плоду и, следовательно, состояние углеводного питания последнего, определяющего внутриутробное развитие [4, 16, 31, 48, 51]. В этом процессе ведущую роль также играют плацентарные гормоны — плацентарный лактоген и хорионический гонадотропин [2]. Плацентарный лактоген обладает широким спектром действия: соматотропным, лактогенным, лютеотроп- ным, а также потенцирует хорионический гонадотропин [35].
При этом циркулирующий в крови матери инсулин не проникает через плаценту, но от его секреции зависит содержание глюкозы в крови матери и поступление к плоду и, следовательно, состояние углеводного питания последнего, определяющего внутриутробное развитие [4, 16, 31, 48, 51]. В этом процессе ведущую роль также играют плацентарные гормоны — плацентарный лактоген и хорионический гонадотропин [2]. Плацентарный лактоген обладает широким спектром действия: соматотропным, лактогенным, лютеотроп- ным, а также потенцирует хорионический гонадотропин [35].
В то же время избыток инсулина может отрицательно влиять на развитие плода. Так, при введении инсулина крысам на 5—7-й день беременности двукратно в течение дня у 12-дневных эмбрионов, извлеченных методом кесарева сечения, обнаруживались пороки развития [20]. Повышение уровня инсулина в амниотической жидкости на ранних стадиях беременности может свидетельствовать о высокой степени риска аномалий развития плода [14].
В наших экспериментах введение крысам на 5—7-й день беременности возрастающих доз инсулина сопровождалось дозозависимым снижением числа родивших животных и количества особей в помете. Вскрытие 10 крыс с 10-дневной беременностью, которым на 5—7-й день после спаривания вводили инсулин из расчета 0,25 ЕД на 100 г массы, показало, что у 2 самок в матке развивалось по 4 эмбриона, а у всех остальных животных этот орган был увеличен и диффузно гиперемирован. В нем отсутствовали признаки имплантации. У 6 из 10 забитых контрольных беременных крыс в обоих рогах матки было обнаружено от 8 до 13 развивающихся эмбрионов, у 4 особей этот орган остался таким же, как у вергинных самок [10].
Вскрытие 10 крыс с 10-дневной беременностью, которым на 5—7-й день после спаривания вводили инсулин из расчета 0,25 ЕД на 100 г массы, показало, что у 2 самок в матке развивалось по 4 эмбриона, а у всех остальных животных этот орган был увеличен и диффузно гиперемирован. В нем отсутствовали признаки имплантации. У 6 из 10 забитых контрольных беременных крыс в обоих рогах матки было обнаружено от 8 до 13 развивающихся эмбрионов, у 4 особей этот орган остался таким же, как у вергинных самок [10].
Следовательно, инсулин, введенный животным в ранние сроки беременности, вызывая гипогликемию, тормозит метаболические процессы в эндометрии матки, играет роль антагониста эстрадиола и прогестерона, которые в ранний внутриутробный период оказывают трофическое влияние на зародыш [4].
Позднее, после образования плаценты, инсулин свое влияние на плод, как отмечалось выше, опосредует через изменения содержания питательных веществ, главным образом глюкозы, в крови матери [33, 43]. Глюкагон, так же как инсулин, включается в регуляцию углеводного обмена. Его секреция активируется при понижении уровня глюкозы в крови. Этот гормон стимулирует фосфорилазу печени, способствует распаду гликогена в ней и выходу глюкозы в кровь. Сведения о его регулирующем влиянии в периоды беременности и внутриутробного развития плода немногочисленны. Установлено, что в развивающейся поджелудочной железе плода глюкагон появляется на 12,5-й день внутриутробного развития [40] и, видимо, играет определенную регуляторную роль уже пренатально. В то же время глюкагон матери не проникает в плод, о чем свидетельствуют опыты на беременных овцах [46]. К концу беременности в крови женщин повышается уровень глюкагона на фоне низких концентраций инсулина и гормона роста [29]. При длительном введении глюкозы самкам крыс в последние дни беременности содержание глюкагона в крови у них снижается при некотором нарастании уровня инсулина |38|. Введение глюкагона извне поздним зародышам вызывает увеличение глюкозо-6-фосфатазной активности и снижение гликогена в печени [25]. Кроме того, показано, что глюкагон, вводимый зародышам крысы, активирует у них ключевой фермент глюконеогенеза — фосфоенолпируваткарбоксикиназу, а также превращение в глюкозу лактата и некоторых аминокислот [28, 34].
Его секреция активируется при понижении уровня глюкозы в крови. Этот гормон стимулирует фосфорилазу печени, способствует распаду гликогена в ней и выходу глюкозы в кровь. Сведения о его регулирующем влиянии в периоды беременности и внутриутробного развития плода немногочисленны. Установлено, что в развивающейся поджелудочной железе плода глюкагон появляется на 12,5-й день внутриутробного развития [40] и, видимо, играет определенную регуляторную роль уже пренатально. В то же время глюкагон матери не проникает в плод, о чем свидетельствуют опыты на беременных овцах [46]. К концу беременности в крови женщин повышается уровень глюкагона на фоне низких концентраций инсулина и гормона роста [29]. При длительном введении глюкозы самкам крыс в последние дни беременности содержание глюкагона в крови у них снижается при некотором нарастании уровня инсулина |38|. Введение глюкагона извне поздним зародышам вызывает увеличение глюкозо-6-фосфатазной активности и снижение гликогена в печени [25]. Кроме того, показано, что глюкагон, вводимый зародышам крысы, активирует у них ключевой фермент глюконеогенеза — фосфоенолпируваткарбоксикиназу, а также превращение в глюкозу лактата и некоторых аминокислот [28, 34]. В поздний зародышевый период повышается отношение инсулин/глюкагон, которое находится в соответствии с гликемическим гомеостазом плода, поддерживаемым за счет поступления глюкозы из материнского организма [54].
В поздний зародышевый период повышается отношение инсулин/глюкагон, которое находится в соответствии с гликемическим гомеостазом плода, поддерживаемым за счет поступления глюкозы из материнского организма [54].
Исследования на млекопитающих животных показали, что одной из функций глюкагона в пренатальном периоде развития является индукция синтеза ферментов печени, в том числе глюкозо-6-фосфатазы, которые участвуют в регуляции обмена глюкозы в период беременности |4].
Таким образом, изложенные выше факты свидетельствуют о серьезных изменениях углеводного обмена, возникающих у животных в период беременности. В самом начале этого периода глюкоза стимулирует процесс имплантации. По мере увеличения сроков беременности в материнском организме усиливается синтез углеводов, но их окисление тканями снижается. Уровень глюкозы в крови беременных животных становится нестабильным и колеблется в широком диапазоне в ответ на различные воздействия. Введенные в полость беременной матки препараты, блокирующие обмен углеводов, а также нарушение кровоснабжения одного из рогов этого органа предотвращают процесс имплантации или развитие эмбрионов. Повышение концентрации глюкозы в крови беременных самок способствует увеличению числа особей в помете и улучшению их жизнеспособности.
Повышение концентрации глюкозы в крови беременных самок способствует увеличению числа особей в помете и улучшению их жизнеспособности.
Согласно данным литературы, после оплодотворения в местах предполагаемой имплантации зародыша повышается потребление кислорода, поэтому вероятней всего глюкоза, сукцинат, пируват, лактат, ацетат повышают в эндометрии матки энергетические проценты и тем самым увеличивают число мест имплантаций и последующее развитие эмбриона [32, 36, 40, 53].
Этот процесс является довольно сложным и до конца не исследованным. Регуляция обмена глюкозы в материн ском организме и стимуляция ростовых процессов плода осуществляются инсулином, инсулиноподобным ростовым фактором I и другими ростовыми факторами. Инсулиноподобный ростовый фактор I стимулирует утилизацию глюкозы, образование лактата, СОг и накопление гликогена в ткани изолированной плаценты человека [22, 54]. В то же время большие дозы инсулина, введенные в ранние сроки беременности, блокируют имплантацию, а позже нарушают процесс внутриутробного развития. Глюкагон также участвует в обмене углеводов у беременной матери и плода. В плане дальнейшего изучения физиологических механизмов обмена углеводов перспективными могут быть исследования, направленные на изучение следующих вопросов:
Глюкагон также участвует в обмене углеводов у беременной матери и плода. В плане дальнейшего изучения физиологических механизмов обмена углеводов перспективными могут быть исследования, направленные на изучение следующих вопросов:
а) молекулярные механизмы действия глюкозы на плод и децидуальную оболочку матки в ранние сроки беременности;
б) механизмы отрицательного действия инсулина в период эмбриогенеза на развивающийся эмбрион;
в) участие глюкагона в регуляции обмена глюкозы в процессе беременности;
г) механизмы взаимоотношения инсулина с основными гормонами беременности (прогестероном и эстрогенами).
1. Алимухамедов А. А. // Съезд физиологов Узбекистана, 4-й: Тезисы науч, сообщений.— Ташкент, 1988.— С. 39.
2. Вельтищев Ю. Е., Ермолаев М. В., Аноненко В. А., Князев Ю. А. Обмен веществ у детей.— М., 1983.
3. Воробьев Н. Н. // Изв. АН СССР. Сер. биол.— 1985,— № 1,— С. 61—70.
4. Држевецкая И. А. Эндокринная система растущего организма.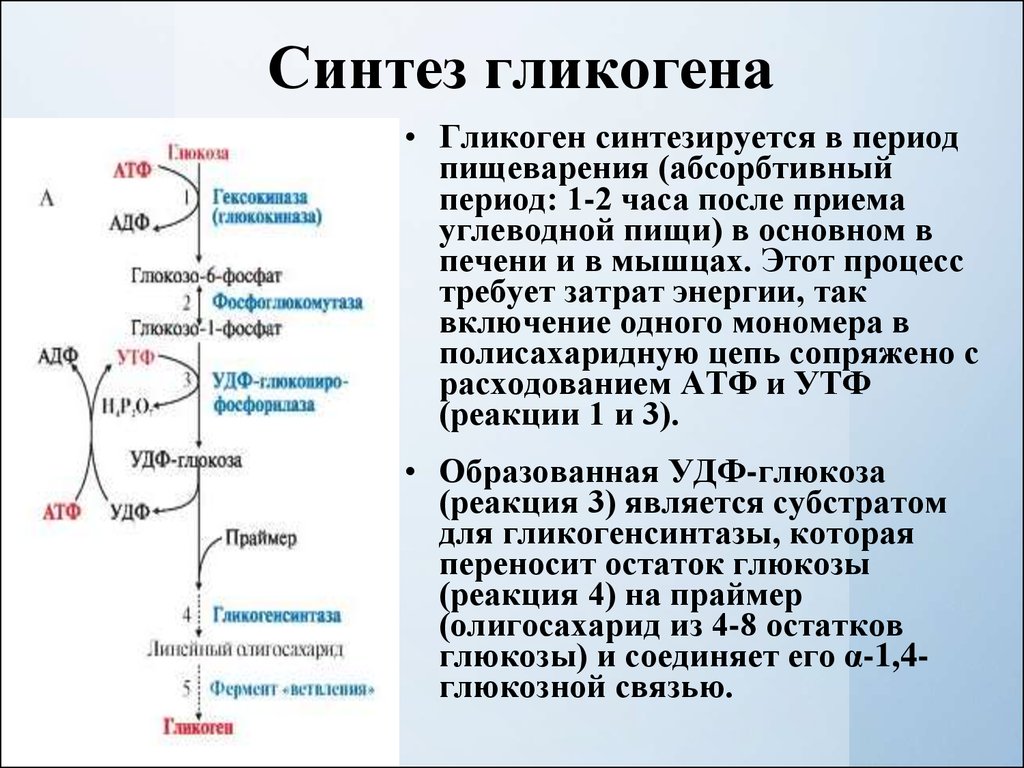 — М., 1987.
— М., 1987.
5. Зыбина Т. Г. // Цитология.— 1983.— Т. 30, № 10.— С. 1180.
6. Каминский Ю. Г., Косенко Е. А. Парадоксы углеводного обмена.— Пущино, 1988.
7. Колесников С. И., Морозова Л. М. Генетико—физиологические взаимоотношения матери и плода.— Новосибирск, 1985.
8. Косенко Е. А., Каминский Ю. Г. Углеводный обмен, печень и алкоголь.— Пущино; 1988.
9. Махмудов Э. С., Алимухамедов А. А., Рахимов К. Р., Садыков Б. А. Рекомендации по снижению яловости коров и сохранению молодняка крупного рогатого скота в условиях Узбекистана. Информ, сообщение № 442.— Ташкент, 1988.
10. Махмудов Э. С., Ахмеров Р. Н., Алимухамедов А. А., Бабаева Р. Н. Реакция беременной самки и ее потомства на введение глюкозы и инсулина. Депонир. в ВИНИТИ 29.03.91 № В 82 — В 71.
11. Рахимов К. Р., Демидова А. И. Углеводы и механизмы их усвоения.— Ташкент, 1986.
12. Репин В. С. // Успехи соврем, биол.— 1976.— Т. 81,— С. 106—125.
13. Репин В. С. Критические факторы химической регуляции развития.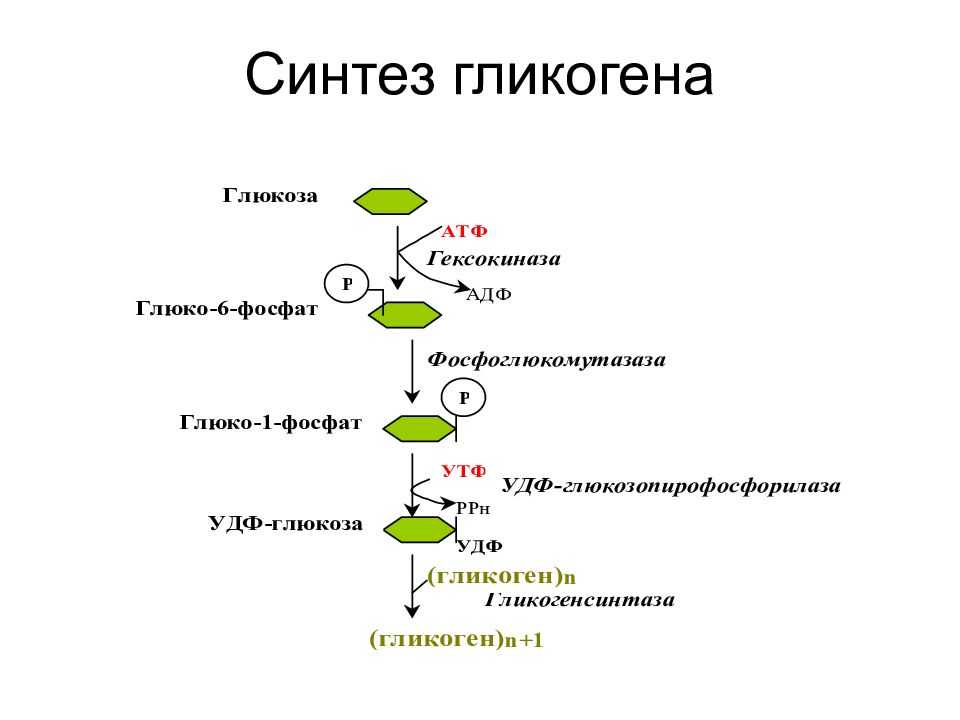 — М., 1980.
— М., 1980.
14. Шевченко Т. К., Абдуллаев Н. X., Шамирзаев Н. X. Беременность и сахарный диабет.— Ташкент, 1988.
15. Юдаев М. А., Афиногенова С. А., Булатов А. А. и др. Цит. по Воробьеву Н. Н. // Изв. АН СССР. Сер. биол.— 1985,— № 1,— С. 61—70.
16. Anand R. S., Languli S., Sperling М. А. // Amer. J. Physiol.— 1980,— Vol. 238, N 6,— Р. 524—532.
17. Barz S., Jeige A., Mitzkat H. J. // Med. Klin.— 1985.— Bd 80, N 18.— S. 483—487.
18. Britta A., Mattson J. V., Rosenblum R. M. et al. .// Diabetes.— 1988,— Vol. 37, N 5,— P. 585—589.
19. Buch L., Hornes P. J., Kuhl C. // Acta endocr. (Kbh.).— 1986,— Vol. 112, N 2 — P. 263—266.
20. Buchanan T. A., Schemmer J. K., Frelnker N. // J. clin. Invest.— 1986,— Vol. 78, N 3.— P. 643—649.
21. Chovez D. J. // Europ. J. Cell Biol.— 1986.— Vol. 42, Suppl. 15.— P. 69.
22. Dieguez C., Page M. D., Peters T. R., Seaulon M. F. // J. roy. Coll. Phycns bond.— 1988.— Vol. 22, N 2,— P. 84—91.
23. Flood M. R„ Wiebold J.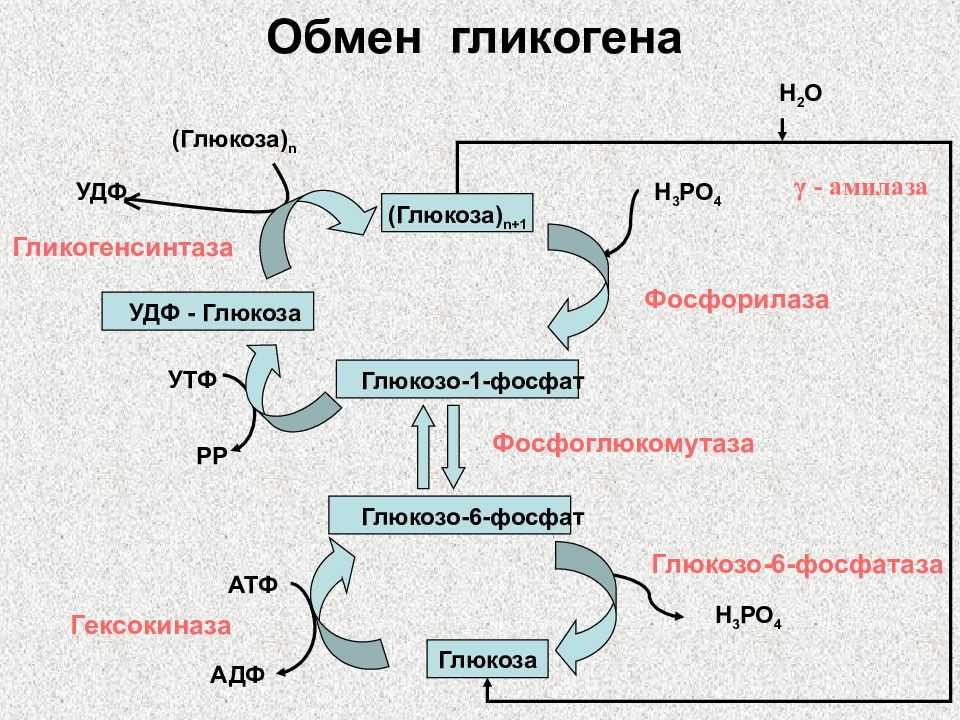 L. // J. Reprod. Fertil.—, 1988,— Vol. 84, N 1.— P. 7—12.
L. // J. Reprod. Fertil.—, 1988,— Vol. 84, N 1.— P. 7—12.
24. Galt A. L., Hardy K., Winston R. M. Z., Leess H. I. // Hum. Reprod.— 1990.— Vol. 5, N I.— P. 104—108.
25. Greengard O., Dowey H. K. // J. biol. Chem.— 1967.— Vol. 242,— P. 2968—2991.
26. Gilbert M., Hauguel S., Bouisset M. // Amer. J. Physiol.— 1981 — Vol. 247, N 5,— P. 574—580.
27. Gilbert M., Sparks 1. W., Girard J., Battaglia J. C. // Biol. Neonat.— 1985,— Vol. 48, N 2.— P. 90—99.
28. Girard J. R., Guillet J., Marty J. et al. // Diabetologia.— 1976,— Vol. 12, N 4,— P. 327—337.
29. Gonzalez- Willamer G., Argota-Espinosa R., Niz-Rames J. // Arch, invest. Med.— 1982,— Vol. 13, N 4,— P. 239— 244.
30. Hainz E. T., Nguyen V. V., Fussgahet R. D. // Biol. Neonate.—1982.— Vol. 41, N 5—6.— P. 240—245.
31. Hay W. W., Sparks J. W., Wilkening R. B. et al. // Amer. J. Physiol.— 1983,— Vol. 245, N 4,— P. E347—E350.
32. Horst C. J. G. // Cytobios.— 1986,— Vol. 45, N 181,— P. 85—95.
33. Itskovitz J., Hodgen G. D. // Psychoneuroendocrino— logy.— 1988,— N 1—2,— P. 155—170.
Itskovitz J., Hodgen G. D. // Psychoneuroendocrino— logy.— 1988,— N 1—2,— P. 155—170.
34. Jeung D., Oliver J. T. // Biochem. J.—1968.— Vol. 108, N 2,— P. 325—327.
35. Josimovich J. B. // Comparative Aspects of Reproductive Failure.— Berlin, 1967.— P. 176—185.
36. Khurana N. K., Wales R. G. // Aust. J. biol. Sci.— 1987 — Vol. 40, N 4,— P. 389—395.
37. Koski K. G„ Hill F. W., Harley L. S. // J. Nutr.— 1986,— Vol. 116, N 10,— P. 1922—1937.
38. Ktorra A., Nurjhan N., Girard J. R., Picon L. // Reprod. Nutr. Develop.— 1983.— Vol. 32, N 2.— P. 332—339.
39. Kozaric Z., Peternal P., Labunzija M. // Vet. Arch.— 1988 —Vol. 58, N 1,—P. 33—39.
40. Leturgue A., Hauguel S., Ferra P., Girard J. // Biol. Neonat.— 1987.— Vol. 51, N 2,— P. 64—69.
41. Leturgue A., Revelli J. P., Hauguel S. et al. // Amer. J. Physiol.— 1987,— Vol. 253, N 6,— Pt 1,— P. 616—620.
42. Manuelle P., Вис H. A., Plas Ch. // Biochim. biophys. Acta. Molec. Cell Res.— 1987,— Vol. 298, N 3,— P.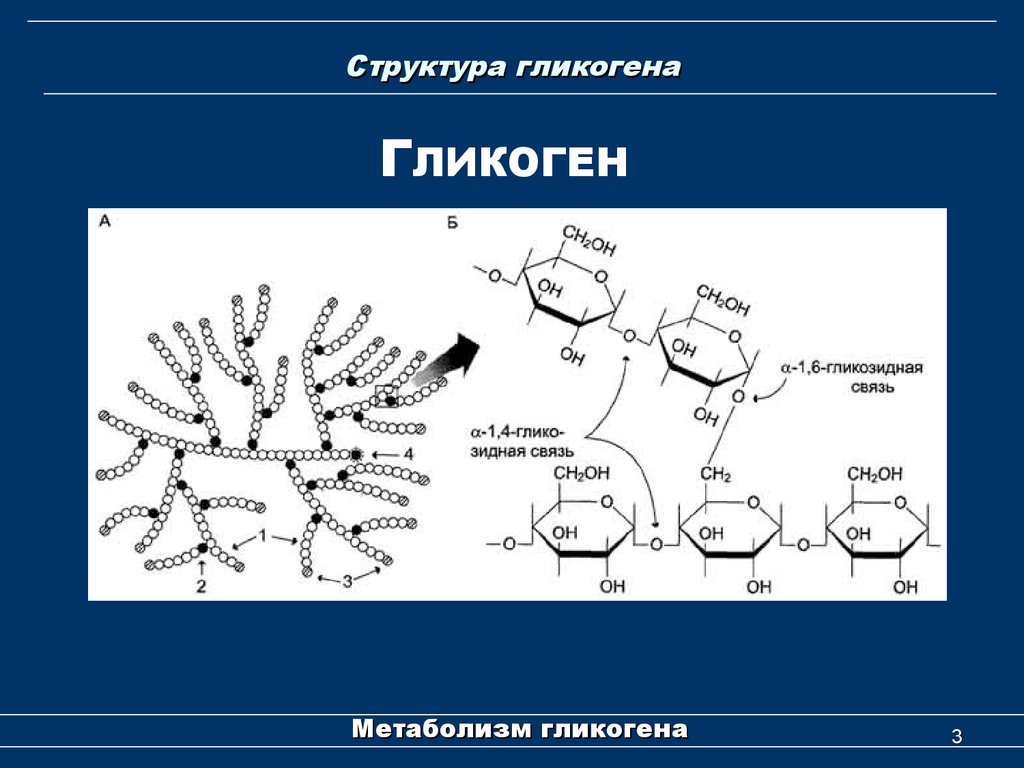 332—340.
332—340.
43. Milner R. D., Hill D. I. // Clin. Endocr.— 1984.— Vol. 21, N 4,— P. 415—433.
44. Naismith D. J., Richardson D. P., Pritchard A. E. // Brit. J. Nutr.— 1982,— Vol. 48.— P. 433—441.
45. Nilsson В. O., Ostensson C. G., Eide S., Hellestrom С. // Endocrinology.— 1980.— Vol. 76, N 1,— P. 82—93.
46. Nitzan M. // Isr. J. med. Sci.— 1981.— Vol. 17, N 5.— P. 378—380.
47. Pinget M., Gander R., Jacques C. et al. // Path Biol.— 1982,— Vol. 30, N 1.— P. 43—48.
48. Rancin J. H. G., Sadarski G., Shanchan M. R. // J. Develop. Physiol.— 1986,— Vol. 8, N 4,— P. 247—253.
49. Singh V. H., Sabnis J. H. // Physiol, and Ecol.— 1986.— Vol. II, N 2,— P. 95—97.
50. Sitianou K., Henrichs J., Teller W. A. // Acta endocr. (Kbh.).— 1988,— Vol. 117, Suppl. 287,— P. 55—56.
51. Sutter-Dub M. T., Dozey B., Vergnaud M. Th., Modes A. M. // Horm. Metab. Res.— 1984.— Vol. 13, N 3.— P. 181 — 184.
52. Toyoda N., Murata K., Sugiama J. // Endocrinology.— 1982 — Vol. 116, N 3,— P. 998—1002.
116, N 3,— P. 998—1002.
53. Vilar R. C., Hicke Z. R. // Arch, invest. Med.— 1988,— Vol. 19, N 3.— P. 283—289.
54. Young A. A., Bogardus E., Stone K., Molt D. M. // Amer. J. Physiol.— 1988,— Vol. 254, N 2,— Pt I.— P. 231—236.
55. Zorzano A., Josuncion M. A., Herrera E. // Metabolism.— 1983 — Vol. 35, N 4,— P. 297—303.
Об авторах
Э. С. Махмудов
Институт физиологии АН Узбекистана
Узбекистан
В. А. Ходжиматов
Институт физиологии АН Узбекистана
Узбекистан
Glycogen Storage Diseases Precision Panel
Болезни накопления гликогена – это группа наследственных заболеваний, вызванных мутациями в генах, которые кодируют ферменты, участвующие в синтезе и расщеплении гликогена, что приводит к его накоплению в тканях организма.

Общая информация
Показания
Клиническое значение
Гены и заболевания
Методология
Ссылки на публикации
Общая информация
- Болезни накопления гликогена – это группа наследственных заболеваний, вызванных мутациями в генах, которые кодируют ферменты, участвующие в синтезе и расщеплении гликогена, что приводит к его накоплению в тканях организма. Существует пять типов болезней накопления гликогена, отличающихся в зависимости от индивидуальной недостаточности ферментов, из которых наиболее распространенным является тип 1 или болезнь фон Гирке. Возраст появления симптомов может варьировать от первых месяцев жизни до более поздних десятилетий жизни. Симптомы возникают из-за нарушения метаболизма гликогена и накопления гликогена в клетках определенных тканей (например, печени и скелетных мышц).
- Панель Igenomix для выявления болезни накопления гликогена может использоваться для постановки направленного диагноза, а также для дифференциальной диагностики гипогликемии, что в конечном итоге приводит к улучшению тактики лечения и прогноза заболевания.
 Исследование обеспечивает всесторонний анализ соответствующих генов с использованием секвенирования нового поколения (NGS).
Исследование обеспечивает всесторонний анализ соответствующих генов с использованием секвенирования нового поколения (NGS).
Показания
- Панель Igenomix для выявления гликогенозов показана пациентам с клиническим подозрением или диагнозом со следующими проявлениями или без них:
- Гипогликемия
- Гепатомегалия
- Задержка рост
- Повышение уровня мочевой кислоты, молочной кислоты и триглицеридов
- Кардиомегалия
- Мышечная гипотония
- Задержка общего моторного развития
- Затрудненное дыхание
- Мышечные судороги и усталость
Клиническое значение
Клиническое значение данной панели:
- Точное молекулярно-генетическое подтверждение клинического диагноза у пациента.
- Раннее начало терапии в виде назначения диеты с частым пероральным приемом глюкозы и заместительной ферментной терапией.

- Оценка статуса носительства и генетическое консультирование бессимптомных членов семьи.
- Точное установление корреляции генотип-фенотип.
Гены и заболевания
Методология
Ссылки
Ссылки на публикации
Ellingwood, S. S., & Cheng, A. (2018). Biochemical and clinical aspects of glycogen storage diseases. The Journal of endocrinology, 238(3), R131–R141. https://doi.org/10.1530/JOE-18-0120
Hicks, J., Wartchow, E., & Mierau, G. (2011). Glycogen storage diseases: a brief review and update on clinical features, genetic abnormalities, pathologic features, and treatment. Ultrastructural pathology, 35(5), 183–196. https://doi.org/10.3109/01913123.2011.601404
Kishnani, P. S., Sun, B., & Koeberl, D. D. (2019). Gene therapy for glycogen storage diseases. Human molecular genetics, 28(R1), R31–R41. https://doi.org/10.1093/hmg/ddz133
S., Sun, B., & Koeberl, D. D. (2019). Gene therapy for glycogen storage diseases. Human molecular genetics, 28(R1), R31–R41. https://doi.org/10.1093/hmg/ddz133
Ross, K. M., Ferrecchia, I. A., Dahlberg, K. R., Dambska, M., Ryan, P. T., & Weinstein, D. A. (2020). Dietary Management of the Glycogen Storage Diseases: Evolution of Treatment and Ongoing Controversies. Advances in nutrition (Bethesda, Md.), 11(2), 439–446. https://doi.org/10.1093/advances/nmz092
Kishnani, P. S., Goldstein, J., Austin, S. L., Arn, P., Bachrach, B., Bali, D. S., Chung, W. K., El-Gharbawy, A., Brown, L. M., Kahler, S., Pendyal, S., Ross, K. M., Tsilianidis, L., Weinstein, D. A., Watson, M. S., & ACMG Work Group on Diagnosis and Management of Glycogen Storage Diseases Type VI and IX (2019). Diagnosis and management of glycogen storage diseases type VI and IX: a clinical practice resource of the American College of Medical Genetics and Genomics (ACMG). Genetics in medicine : official journal of the American College of Medical Genetics, 21(4), 772–789. https://doi.org/10.1038/s41436-018-0364-2
Genetics in medicine : official journal of the American College of Medical Genetics, 21(4), 772–789. https://doi.org/10.1038/s41436-018-0364-2
Derks, T., Oosterveer, M. H., & De Souza, C. F. (2018). Next-generation glycogen storage diseases. Journal of inherited metabolic disease, 41(6), 911–912. https://doi.org/10.1007/s10545-018-00250-0
descargarПодробное описание
Загрузить
БРОШЮРА
Загрузить
Чем мы можем вам помочь?
-
Имя
-
Фамилия
-
Email:
-
Телефон
-
Должность Пациент ЭКО Пациент-акушер-гинеколог GPDx Patient General OBGYN Сонографист Медсестра Педиатр Неврология Кардиология Клиническая генетика Гематология и гемотерапия Иммунология Аллергология Гастроэнтерология Дерматология и венерология Эндокринология и питание Клиническая фармакология Внутренняя медицина Нефрология Пневмология Клиническая нейрофизиология Офтальмология Медицинская онкология Оториноларингология Психиатрия Ревматология Урология Врач ЭКО Эмбриолог Главный эмбриолог IVF Geneticist Медицинский директор Другое
-
Услуга: про которую я хотел(а) бы узнать Genomic Precision DiagnosticGenetic Counseling
-
Accept the privacy policy
reCAPTCHA demo: Simple page
ГЛИКОГЕН цена, купить ГЛИКОГЕН — химическая книга
ГЛИКОГЕН
- $44 — $3250
- Название продукта: ГЛИКОГЕН
- КАС: 9005-79-2
- МФ: (C6h20O5)n
- МВт: 666,58
- EINECS: 232-683-8
- Лей Номер: MFCD00081547
- Синонимы: Гликоген(Ⅲ) из печени кролика;(2S,3R,4S,5S,6R)-2-[[(2R,3S,4R,5R,6R)-4,5-дигидрокси-3-[ (2R,3R,4S,5S,6R)-3,4,5-тригидрокси-6-(гидроксиметил)оксан-2-ил]окси-6-[(2R,3S,4R,5R,6S)-4, 5,6-тригидрокси-2-(гидроксиметил)оксан-3-ил]оксиоксан-2-ил]метокси]-6-(гидроксиметил)оксан-3,4,5-триол; Гликоген из устриц Vetec™ реактивной квалификации , Тип II; Гликоген из устриц Тип II; Раствор гликогена (20 мг/мл) из устриц, протестирован на нуклеазу; животный крахмал; Гликоген Тип IX; лиогликоген
43 цены
Выбранное состояние:
Марка
- АК Научный
- Альфа Аесар
- Биосинт Карбосинт
- Химсцена
- Специализированные химикаты Frontier
- Матрица Научная
- Сигма-Олдрич
- Усбиологический
Пакет
- 25 мг
- 50 мг
- 100 мг
- 250 ул
- 250 мг
- 500 мг
- 1 мл
- 1 г
- 5г
- 10 г
- 25 г
- 50 г
- 1 флакон
- 100 г
- 250 г
- 500 г
- ПроизводительAK Scientific
- Номер продуктаM540
- Описание продуктаГликоген
- Упаковка500 г
- Цена $1539
- Обновлено 16.
 12.2021
12.2021 - Купить
- ПроизводительAlfa Aesar
- Номер продуктаJ16445
- Описание продуктаГликоген из устриц, ультрачистый, Affymetrix/USB
- Упаковка5г
- Цена $75,91
- Обновлено 06.01.2023
- Купить
- ПроизводительAlfa Aesar
- Номер продуктаJ16445
- Описание продуктаГликоген из устриц, ультрачистый, Affymetrix/USB
- Упаковка 25 г
- Цена $332,05
- Обновлено 06.01.2023
- Купить
- ПроизводительAlfa Aesar
- Номер продуктаJ16445
- Описание продуктаГликоген из устриц, ультрачистый, Affymetrix/USB
- Упаковка 100 г
- Цена $1282
- Обновлено 06.
 01.2023
01.2023 - Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG71459
- Описание продуктаГликоген из печени кролика
- Упаковка50 мг
- Цена $75
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продуктаYG162526
- Описание продуктаГликоген из бычьей печени
- Упаковка 250 мг
- Цена $100
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG71459
- Описание продуктаГликоген из печени кролика
- Упаковка 100 мг
- Цена $120
- Обновлено 16.
 12.2021
12.2021 - Купить
- ПроизводительBiosynth Carbosynth
- Номер продуктаYG162526
- Описание продуктаГликоген из бычьей печени
- Упаковка500 мг
- Цена $150
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG71459
- Описание продуктаГликоген из печени кролика
- Упаковка 250 мг
- Цена $200
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продуктаYG162526
- Описание продуктаГликоген из бычьей печени
- Упаковка 1 г
- Цена $250
- Обновлено 16.
 12.2021
12.2021 - Купить
- ПроизводительBiosynth Carbosynth
- Номер продуктаYG71459
- Описание продуктаГликоген из печени кролика
- Упаковка5G
- Цена $1200
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG40133
- Описание продуктаГликоген из устриц
- Упаковка 100 г
- Цена $1450
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG71459
- Описание продуктаГликоген из печени кролика
- Упаковка10 г
- Цена $2100
- Обновлено 16.
 12.2021
12.2021 - Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG40133
- Описание продуктаГликоген — из устрицы
- Упаковка 250 г
- Цена $3250
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG40133
- Описание продуктаГликоген из устриц
- Упаковка10 г
- Цена $250
- Обновлено 16.12.2021
- Купить
- ПроизводительBiosynth Carbosynth
- Номер продукта YG71459
- Описание продуктаГликоген из печени кролика
- Упаковка500 мг
- Цена $300
- Обновлено 16.
 12.2021
12.2021 - Купить
| Производитель | Номер продукта | Описание продукта | Упаковка | Цена | Обновлено | Купить |
|---|---|---|---|---|---|---|
| АК Научный | М540 | Гликоген | 500 г | 1539 $ | 2021-12-16 | Купить |
| Альфа Аесар | J16445 | Гликоген, из устриц, Ultrapure, Affymetrix/USB | 5 г | 75,91 $ | 06. 01.2023 01.2023 | Купить |
| Альфа Аесар | Дж16445 | Гликоген из устриц, ультрачистый, Affymetrix/USB | 25 г | 332,05 $ | 06.01.2023 | Купить |
| Альфа Аесар | Дж16445 | Гликоген, из устриц, Ultrapure, Affymetrix/USB | 100 г | 1282 $ | 06.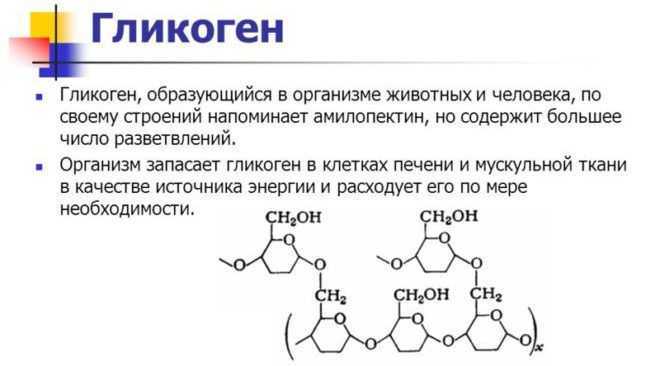 01.2023 01.2023 | Купить |
| Биосинт Карбосинт | ИГ71459 | Гликоген — из печени кролика | 50 мг | 75 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ162526 | Гликоген из бычьей печени | 250 мг | 100 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ71459 | Гликоген — из печени кролика | 100 мг | 120 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ162526 | Гликоген из бычьей печени | 500 мг | 150 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ71459 | Гликоген — из печени кролика | 250 мг | 200 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ162526 | Гликоген из бычьей печени | 1 г | 250 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | YG71459 | Гликоген — из печени кролика | 5G | 1200 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ40133 | Гликоген — из устриц | 100 г | 1450 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ71459 | Гликоген — из печени кролика | 10 г | 2100 $ | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ40133 | Гликоген — из устриц | 250 г | 3250 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ40133 | Гликоген — из устриц | 10 г | 250 долларов | 2021-12-16 | Купить |
| Биосинт Карбосинт | ИГ71459 | Гликоген — из печени кролика | 500 мг | 300 долларов | 2021-12-16 | Купить |
- 1
- 2
- 3
СледующийПоследний
Описание
Гликоген устриц использовался для индукции воспаления и стимуляции экстравазации гранулоцитов и нейтрофилов на животных моделях после внутрибрюшинной инъекции. Гликоген также действует как проба сосудистой проницаемости.
Гликоген также действует как проба сосудистой проницаемости.Цены на сопутствующие товары
ЭтилизоцианоацетатТЕРТ-БУТИЛИЗОЦИАНИДТозилметилизоцианидN-БУТИЛИЗОЦИАНИДМЕТИЛИЗОЦИАНОАЦЕТАТ
Цена на соответствующий продукт
- КОБАТАТ 8 $43-501,45
- Ацетилацетонат железа
$6-2359,48 - Ацетилацетонат меди
$6-826
Поставщики и производители
Хэбэй Guanlang биотехнологии Co., Ltd. 10,00 $ /1 ШТ.
ГЛИКОГЕН
КАС:9005-79-2
 , Ltd.
BOC наук
Чжэнчжоу Альфа Химическая Компания, ООО
Карьера Henan Chemica Co.
Hefei TNJ Chemical Industry Co., Ltd.
Дидеу Индастриз Групп Лимитед
АФАЙН КЕМИКАЛС ЛИМИТЕД
Файнтек Индастри Лимитед
, Ltd.
BOC наук
Чжэнчжоу Альфа Химическая Компания, ООО
Карьера Henan Chemica Co.
Hefei TNJ Chemical Industry Co., Ltd.
Дидеу Индастриз Групп Лимитед
АФАЙН КЕМИКАЛС ЛИМИТЕД
Файнтек Индастри ЛимитедМышечный гликоген — On Your Mark Nutrition
Мышцы
Гликоген
Проверьте восстановление внутри запасов топлива в мышцах
Готов ли я к выступлению?
Достаточно ли моего восстановления?
Насколько полон мой «бак»?
Сколько топлива я использую, когда тренируюсь/тренируюсь/соревнуюсь?
Насколько хорошо заживает травма мягких тканей? Как скоро я смогу вернуться к «нормальному» состоянию?
Есть ли у меня сбалансированное развитие мышц? Как это могло повлиять на меня на поле и за его пределами?
Вышеуказанные вопросы могут быть решены с помощью ультразвуковой технологии MuscleSound®, которая оценивает состояние мышечной энергии (MES) — источник производства мышечной энергии, который состоит из мышечного гликогена и мышечной жидкости. Мышечный гликоген — это место, где хранится большая часть углеводов. Если в мышцах мало гликогена, это может привести к неспособности работать на максимальном уровне и/или повышенному риску получения травмы.
Мышечный гликоген — это место, где хранится большая часть углеводов. Если в мышцах мало гликогена, это может привести к неспособности работать на максимальном уровне и/или повышенному риску получения травмы.
Тестирование мышечного гликогена — еще один показатель в наборе инструментов для оценки восстановления (имеется в виду либо питание, и/или отдых, и/или изменение тренировочной нагрузки) на основе тренировочной нагрузки спортсмена. Должны ли они тренироваться или им следует больше сосредоточиться на каком-то аспекте восстановления? Если они тренируются, могут ли они подвергаться повышенному риску получения травмы из-за дисбаланса гликогена в мышцах справа и слева? Есть ли гиперактивация ягодичных мышц, чтобы компенсировать плохую работу квадрицепсов?
Почему важно измерять уровень гликогена в мышцах?
Готовность : убедиться, что вы готовы к тренировке или соревнованию.
 Важно сканировать как минимум за 2 дня, чтобы увидеть ваши значения. Если они низкие, то ваши мышцы не находятся в оптимальной готовности к работе. Это служит «ранним предупреждением», чтобы вы могли скорректировать свой план питания и/или тренировочную нагрузку.
Важно сканировать как минимум за 2 дня, чтобы увидеть ваши значения. Если они низкие, то ваши мышцы не находятся в оптимальной готовности к работе. Это служит «ранним предупреждением», чтобы вы могли скорректировать свой план питания и/или тренировочную нагрузку.Восстановление : чтобы обеспечить достаточное восстановление для будущих тренировок или соревнований. Важно провести сканирование через 2 дня после тренировки/соревнования, чтобы мышцы успели восстановиться и вернуться к исходному уровню. Если ваше значение низкое, то для восстановления потребуется либо больше отдыха, либо изменения в питании/тренировках.
Отслеживание топлива : сканирование мышц до и сразу после тренировки/соревнования для оценки влияния на мышечный гликоген путем наблюдения за снижением значений MES до и после.
Травма Восстановление :
Если спортсмен все еще восстанавливается после травмы, он по-прежнему предпочитает определенную сторону? Они тебе это скажут? Или даже знать это? Может ли знание этого предотвратить дальнейшие травмы?
Поврежденные мышцы имеют меньшую способность накапливать и использовать топливо по сравнению со здоровыми мышцами.
 Это влияет на ваш энергетический статус мышц. По мере того, как вы проходите реабилитацию, способность накапливать топливо увеличивается, когда ваши мышцы начинают восстанавливаться. Мониторинг и отслеживание улучшений — это инструмент для оценки процесса восстановления. Это избавляет от догадок при лечении, что может ускорить возвращение в игру.
Это влияет на ваш энергетический статус мышц. По мере того, как вы проходите реабилитацию, способность накапливать топливо увеличивается, когда ваши мышцы начинают восстанавливаться. Мониторинг и отслеживание улучшений — это инструмент для оценки процесса восстановления. Это избавляет от догадок при лечении, что может ускорить возвращение в игру.
Мышцы Топливо Симметрия :
Мышечный баланс (двусторонняя мышечная симметрия) является ключом к эффективному движению и хорошей осанке. При упражнениях высокой интенсивности это служит риском получения травмы, особенно во время соревнований. Симметрия мышц между правой и левой сторонами тела помогает предотвратить травмы и потенциально может позволить телу работать более эффективно.
Это новая, очень точная мера симметрии мышечного топлива по сравнению с тестами на симметрию, включающими рудиментарные методы, такие как гимнастика или тестирование силы.

