Тема 7. Общее понятие об обмене веществ и энергии в организме — Студопедия
План
1. Понятие «обмен веществ и энергии», «анаболизм», катаболизм. Взаимосвязь процессов анаболизма и катаболизма. Понятие о промежуточном, внешнем, пластическом и функциональном обменах. Возрастные изменения обмена веществ.
2. Основные этапы преобразования энергии в организме. Окисление – основной путь освобождения энергии. Типы окислительных реакций в организме: прямое присоединение кислорода, отщепление водорода, перенос электронов. Понятие об аэробном и анаэробном биологическом окислении.
3. Макроэргические связи, накопление энергии биологического окисления в макроэргические соединения, их роль в организме. Особая роль АТФ в энергетическом обмене.
1-й вопрос
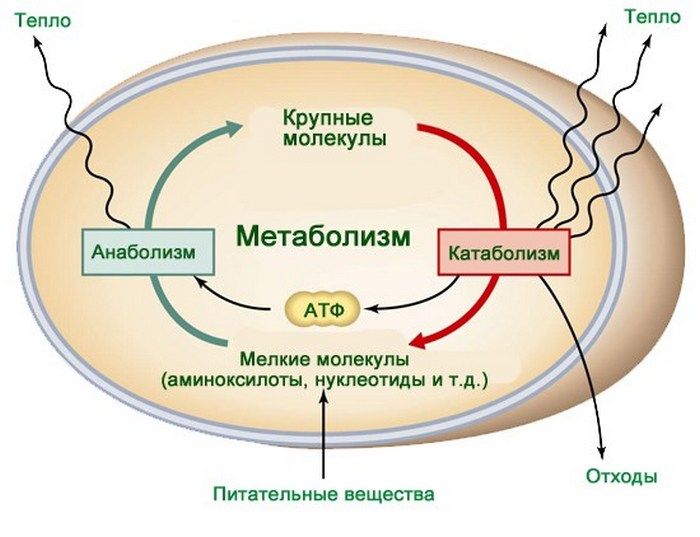
Жизнь – особая форма существования и движения материи, принципиально отличная от других форм. Одним из признаков живой материи является обмен веществ и энергии.
Обмен веществ – это непрерывный, саморегулируемый, самосовершающийся круговорот веществ, протекающий в живой материи и сопровождающийся ее постоянным самообновлением.
Обмен веществ – сложная цепь биохимических реакций, заключающаяся в усвоении веществ из окружающей среды, сложных превращениях их в организме и выделении в окружающую среду продуктов расщепления этих веществ.
Обмен веществ представляет собой сочетание многих разнообразных процессов, среди которых выделяют: катаболизм (диссимиляция), анаболизм (ассимиляция).
Катаболизм – это совокупность процессов распада веществ, составляющих живой организм, распад элементов живого тела на более простые вещества, выделяемые в окружающую среду как конечные продукты жизнедеятельности. Такими конечными веществами являются углекислый газ, вода, аммиак, мочевина и др.; сопровождается выделением энергии.
Анаболизм – это процессы синтеза сложных молекул из более простых, сопровождающиеся поглощением энергии.
Катаболизм и анаболизм – это две теснейшим образом взаимосвязанные стороны процесса обмена веществ. Катаболизм сопровождается освобождением энергии, которая аккумулируется в виде молекул АТФ. При анаболических процессах происходит потребление энергии, которая освобождается при распаде АТФ до АДФ и фосфорной кислоты или АМФ и пирофосфорной кислоты. Таким образом, АТФ является сопрягающим энергетическим звеном катаболизма и анаболизма. Кроме АТФ связующим звеном могут служить специфические метаболические пути или циклы, примером которого является цикл Кребса. Такие пути называют амфиболические.
При анаболических процессах происходит потребление энергии, которая освобождается при распаде АТФ до АДФ и фосфорной кислоты или АМФ и пирофосфорной кислоты. Таким образом, АТФ является сопрягающим энергетическим звеном катаболизма и анаболизма. Кроме АТФ связующим звеном могут служить специфические метаболические пути или циклы, примером которого является цикл Кребса. Такие пути называют амфиболические.
Взаимосвязь катаболизма и анаболизма показана на схеме:
Белки Углеводы Липиды
АДФ + Н3РО4
АТФ
Продукты Продукты
катаболических анаболических
процессов процессов
Цикл Кребса
СО2 Н2О
В обмене веществ принято выделять:
Внешний обмен – это внеклеточное превращение веществ на путях поступления и выделения. В качестве примера можно привести процессы гидролиза углеводов, белков, липидов в желудочно-кишечном тракте.
Промежуточный обмен, или метаболизм – это процессы преобразования компонентов пищи после их переваривания и всасывания (т.е. индивидуальных химических соединений) при их распаде и синтезе на уровне клетки в процессе жизнедеятельности организма. Вещества, образующиеся в ходе химических реакций, называются метаболитами.
Пластический обмен – комплекс химических реакций, приводящих к синтезу специфических для организма веществ: структурных веществ, ферментов, гормонов, различных секретов, запасных источников энергии.
Функциональный обмен – это комплекс реакций, обеспечивающих функциональную активность клетки, органа, ткани. Например, реакции, обеспечивающие мышечное сокращение, работу сердца, печени, почек, легких.
Энергетический обмен организма – это комплекс химических реакций, в процессе которых за счет энергии, освобождающейся при распаде углеводов, жиров, продуктов белкового обмена, происходит ресинтез (новообразование) АТФ, распавшихся в процессе энергетического обеспечения функциональной или пластической деятельности клеток.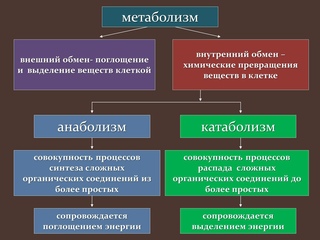
Между пластическим и функциональным обменами существуют конкурентные отношения за обладание определенными веществами, например, АТФ. Если клетки проявляют функциональную активность, поток энергии направляется на ее обеспечение, реакции пластического обмена в это время заметно угнетаются из-за дефицита энергии. Так, при напряженной мышечной работе в организме резко замедляются все процессы синтеза, за исключением синтеза некоторых гормонов, некоторого количества углеводов. после прекращения или снижения функциональной активности процессы пластического обмена усиливаются.
Соотношение процессов катаболизма и анаболизма, скорость протекания изменяются как на протяжении жизни организма, так и в течение определенных периодов времени.
Молодой растущий организм характеризуется заметным преобладание синтеза веществ над распадом. За счет синтеза структурных белков, ферментов, липидов обеспечивается деление клеток и, как следствие, рост организма, увеличение объема тканей и органов.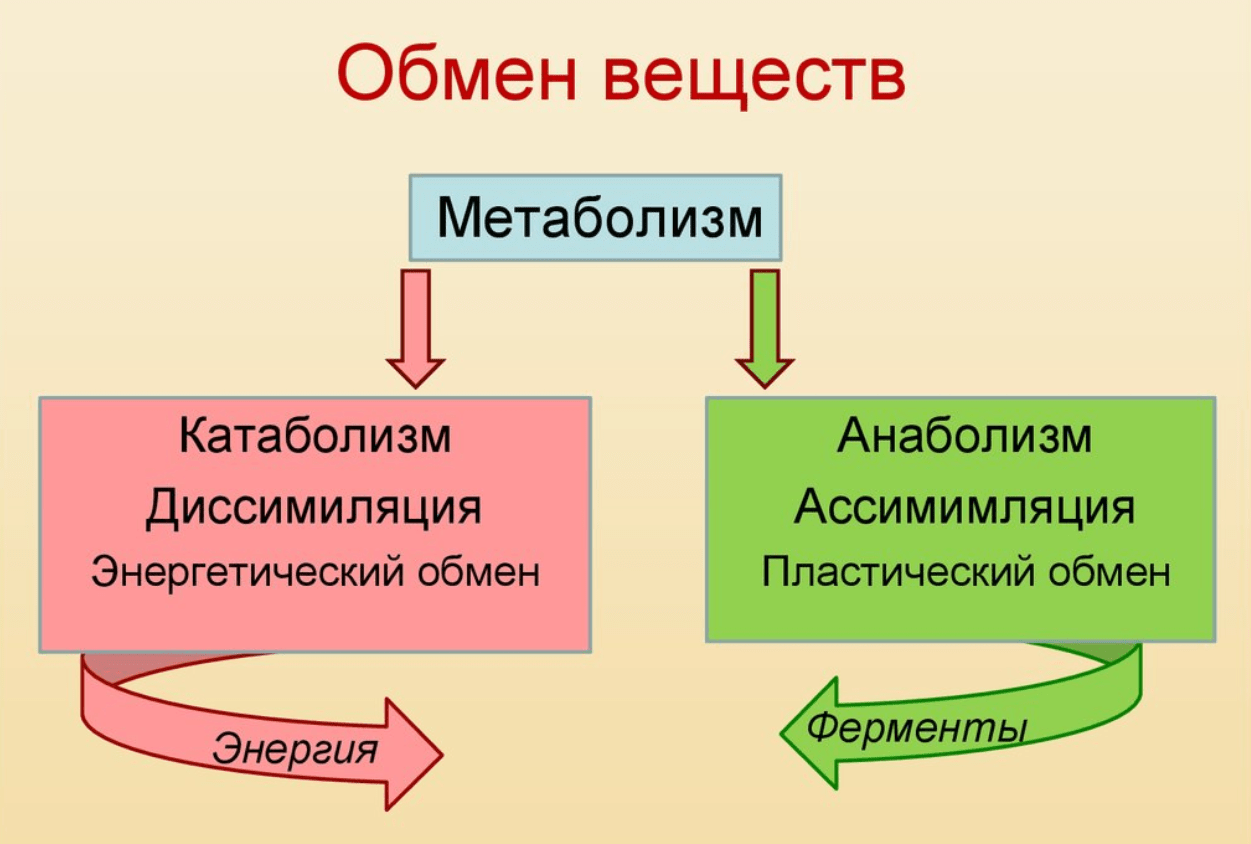
К 17-19 годам в организме устанавливается динамическое равновесие между двумя сторонами обмена веществ. К старости начинают преобладать процессы катаболизма, что приводит к уменьшению содержания важнейших для жизнедеятельности веществ, уменьшению количества клеток в важнейших органах и тканях (головном мозгу, сердце, внутренних органах, мышцах). Данные изменения приводят к снижению функциональных возможностей органов, деятельности организма в целом.
Процессы жизнедеятельности связаны с постоянными затратами энергии. Источником энергии для всех видов биологической работы служит потенциальная химическая энергия, заключенная в молекулах пищевых веществ, она высвобождается в процессе обмена веществ и при помощи специальных молекулярных устройств преобразуется в различные другие виды энергии. В живом организме потенциальная энергия представлена в форме химической энергии связей между атомами в молекулах биоогранических соединений. Например, количество потенциальной энергии, заключенной в молекуле глюкозы в связях между атомами С, Н и О, составляет около 285 кДж/моль вещества.
2-й вопрос
Чтобы высвободить энергию, заключенную в молекулах веществ пищи, последние должны быть подвергнуты ряду специфических превращений в процессе метаболизма.
Процессы преобразования энергии в живых организмах подчиняются законам термодинамики (раздел физики). Который для рассмотрения данных вопросов использует величины: энтальпия, энтропия, кинетическая энергия, свободная энергия. Условно, преобразование энергии можно трактовать следующим образом:
Свободная энергия – та часть энергии системы, которая может быть использована на выполнение работы.
В совокупности всех метаболических реакций, связанных с преобразованием энергии в организме следует выделять:
1. Первичную энергопоставляющую реакцию, где наблюдается небольшое изменение свободной энергии (такие реакции выполняют роль биологического генератора энергии).
2. Реакцию или серию реакций, где выделившаяся свободная энергия связывается в промежуточном соединении, способном к дальнейшему переносу энергии ( энергетическое сопряжение).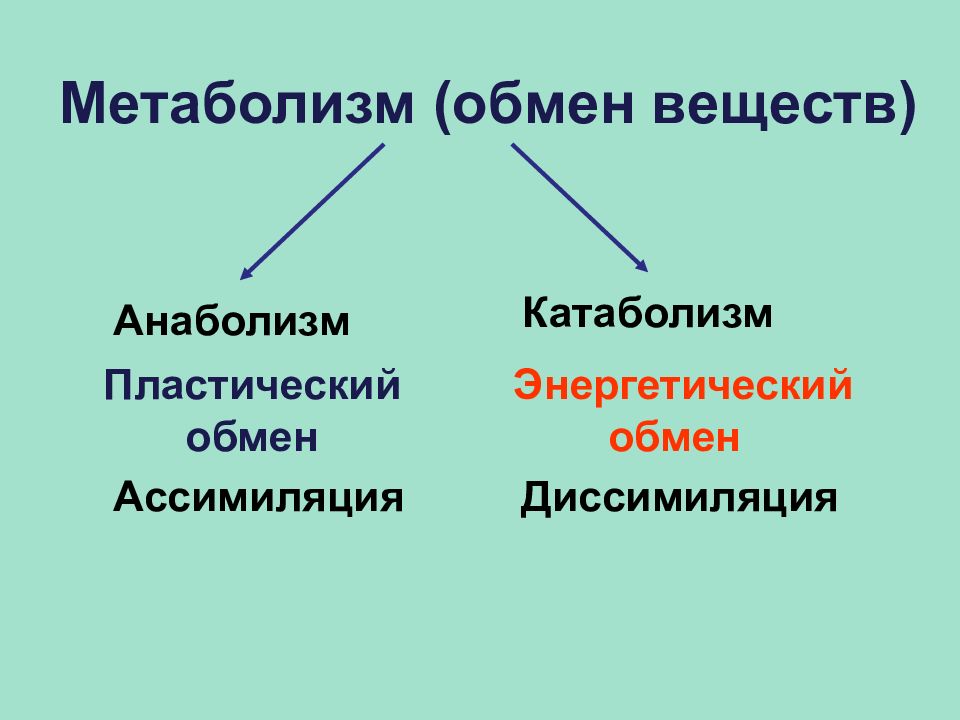
3. Реакцию синтеза стабильного макроэргического соединения, аккумулирующего свободную энергию, которая освобождается в ходе метаболических превращений (в большинстве случаев такой аккумулятор АТФ).
4. Реакции, связанные с использованием энергии макроэргических соединений для синтеза сложных биоорганических соединений и для выполнения различного рода биологической работы.
| Окисление углеводов, жиров, белков | 1,3-дифосфоглицериновая кислота. Фосфоенолпиро-виноградная кислота | АТФ КрФ | Биосинтез белков, жиров, углеводов и др. орг веществ Секреция Мышечное сокращение | |||
| Энергопоставляющие реакции | Энергетическое сопряжение | Аккумуляция энергии | Использование энергии |
Освобождение энергии связано с окислительными процессами. А.Л. Лавуазье в Х VIII веке установил, что при горении органических веществ происходит окисление, т.е. соединение углерода и водорода с кислородом с образованием углекислого газа и воды. В организме окисление органических веществ отличается от процессов горения по своему химизму и условиям протекания (сравнительно низкой температуре тела в присутствии воды, которая вне организма препятствует горению). Процесс получил название тканевое дыхание.
А.Л. Лавуазье в Х VIII веке установил, что при горении органических веществ происходит окисление, т.е. соединение углерода и водорода с кислородом с образованием углекислого газа и воды. В организме окисление органических веществ отличается от процессов горения по своему химизму и условиям протекания (сравнительно низкой температуре тела в присутствии воды, которая вне организма препятствует горению). Процесс получил название тканевое дыхание.
Окислительные реакции связаны с перераспределением электронов с внешних слоев атомов или ионов. Процесс отдачи электронов трактуется как процесс окисления и частица (атом, ион) является донатором электронов, окисляемым веществом. Вещества, принимающие электроны, являются акцепторами электронов, восстанавливаемым веществом. Процесс присоединения электронов называется восстановлением. Окислительные реакции являются окислительно-восстановительными, для их осуществления всегда необходимо наличие системы донатор-акцептор.
При отсутствии какого-либо компонента системы окислительные процессы невозможны.
Окислительные процессы в организме протекают несколькими типами:
1. Присоединение кислорода.
Происходит при образовании воды из водорода и кислорода. В результате взаимодействия двух атомов Н, отдающих электроны (окисляются) атому кислорода (восстанавливается):
2е
2 Н° + О° 2Н + + О2– Н2О + 56, 7 ккал
Н
׀ О
СН3 – С – ОН + ½ О СН3 – С – Н + Н2О
׀
Н
Вдыхаемый организмом кислород, используется как акцептор водорода и входит в состав образующейся воды. Углекислый газ образуется за счет тех атомов кислорода, которые входят в состав окисляемого вещества.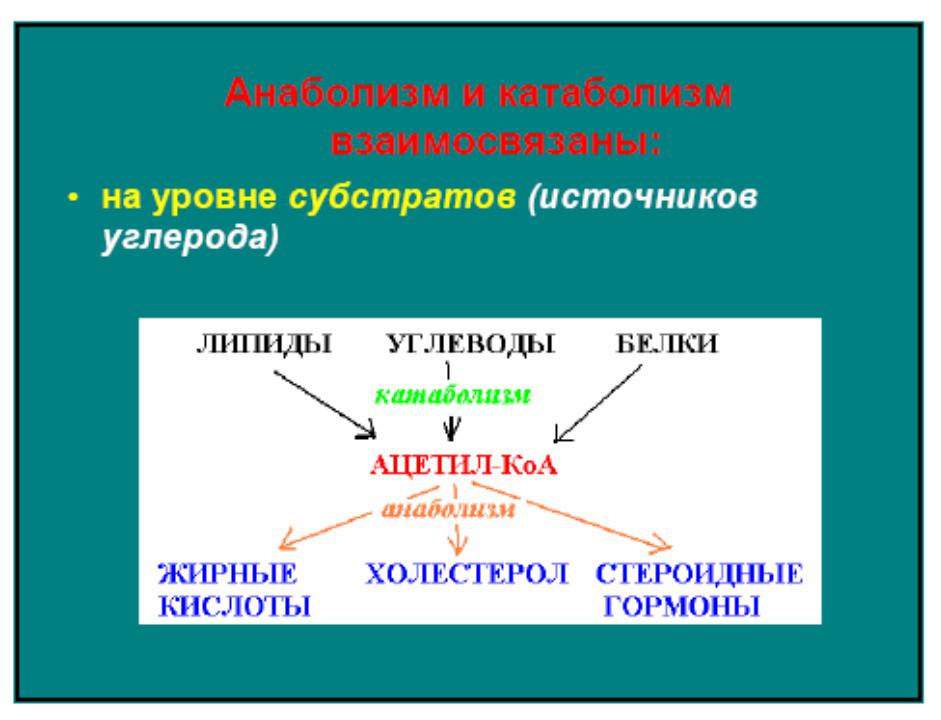 Однако, даже в наиболее богатых кислородом органических веществ – углеводы – кислорода содержится меньше, чем в образующихся конечных продуктах окисления глюкозы:
Однако, даже в наиболее богатых кислородом органических веществ – углеводы – кислорода содержится меньше, чем в образующихся конечных продуктах окисления глюкозы:
С6 Н12 О6 + 6О2 6 СО2 + 6 Н2О
Видно, что для образования 6 молекул СО2 необходимы атомы О.
Присоединение кислорода происходит не в виде молекулярного, а виде воды (она в уравнении не показана).
2. Перенос электронов.
Взаимодействие водорода с кислородом осуществляется благодаря функционированию ферментной системы: цепи переносчиков. Ферменты, катализирующие отнятие Н (дегидрогеназы), ферменты, катализирующие присоединение Н к О (оксидазы), имеют белковую природу и сложное строение.
При транспортировки атома Н по цепи переносчиков происходит освобождение Q. Причем некоторые ферменты могут переносить атом Н (Н+ – протон, ядро; ē – электрон), другие – только ē.
В виде схемы это можно показать так:
H+
Н (Н+е—) Н+е— 2е— 2е— 2е— 2е— 2е—
S НАД ФАД b c a1 a3 1/2O2 H2O
Н (Н+е—) Н+е—
H+
Процессы окисления могут происходить как с участием кислорода (аэробно), так и без его участия (анаэробно).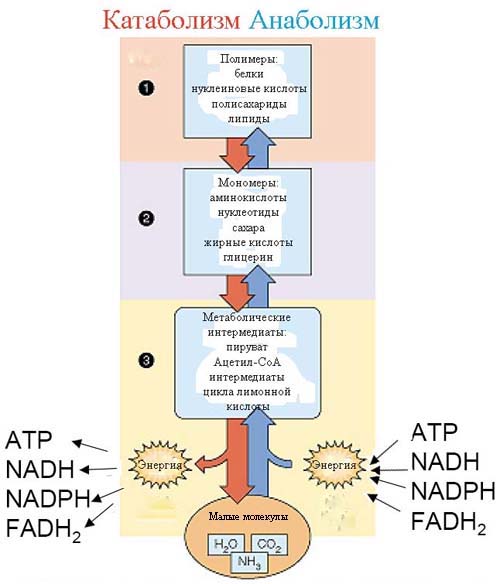 Существует немало организмов (многие бактерии, паразитические черви и др.), которые черпают энергию только из процессов анаэробного окисления и гибнут в присутствии кислорода.
Существует немало организмов (многие бактерии, паразитические черви и др.), которые черпают энергию только из процессов анаэробного окисления и гибнут в присутствии кислорода.
У большинства животных и человека преобладают процессы аэробного окисления. Без кислорода человек жить не может. Однако, многие окислительное реакции в организме человека могут протекать анаэробно (гликолиз – анаэробное окисление углеводов в мышцах). Данный процесс резко усиливается при интенсивной мышечной деятельности и способствует выполнению работы мышц при недостатке снабжения кислородом.
3-й вопрос
Окислительные процессы сопровождаются освобождением энергии, которая запасается в макроэргических связях. Макроэргическими веществами являются:
а) NН О
׀׀ ׀׀
СООН – СН2 – N – С – N – О ~ Р – ОН Креатинфосфат ( КрФ)
׀ ׀ ׀
Н СН3 ОН
б) О О
׀׀ ׀׀
С – О ~ Р – ОН
׀ ׀
СН – ОН ОН 1,3 — дифосфоглицериновая
׀
С Н2 – О ~ Р – ОН кислота
/׀׀
ОН О
О
в) //
СН3 – С ~ ацетилфосфат
г)
О
׀׀
С – ОН ОН
׀ ׀
С Н – О ~ Р – ОН фосфоэнолпировиноградная кислота (фосфо-ен ПВК)
׀ ׀׀
СН2 ОН О
д) АДФ и АТФ аденозинфосфаты
Различаются нуклеотиды по количеству Ф(н): НМФ (нуклеозидмонофосфаты), НДФ (нуклеозиддифосфаты), НТФ(нуклеозидтрифосфаты.
| O | O | O | ||||||||||||||||||||||||||||||||
| || | || | || | ||||||||||||||||||||||||||||||||
| CH2 | — | O | — | P | — | O | ~ | P | — | O | ~ | P | — | OH | ||||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||||||||||
| OH | OH | OH |
Формула АТФ
Нуклеозидтрифосфаты (НТФ) – самые распространенные макроэргические соединения живой клетки. За счет реакций взаимного перехода АДФ и Ф(н) образуется АТФ и Н2О. Процесс идет при поглощении энергии и фермента:
За счет реакций взаимного перехода АДФ и Ф(н) образуется АТФ и Н2О. Процесс идет при поглощении энергии и фермента:
АДФ + Ф(н) АТФ + Н2О
Обратный процесс идет с разрывом макроэргической связи, и в клетку выходит энергия.
Нуклеозидтрифосфаты (НТФ) – сырье для биосинтеза нуклеиновых кислот.
Нуклеозидмонофосфаты (НМФ) – структурные звенья нуклеиновых кислот.
Нуклеозидмонофосфаты (НМФ) входят в состав ферментов (это коферменты ДКФ).
Нуклеозиддифосфаты (НДФ) – участники синтеза ди- и полисахарадов.
АТФ играет наиболее важную роль в энергетическом обмене. Молекула АТФ состоит из органического основания аденина, углевода рибозы и трех остатков фосфорной кислоты, связи между первым и вторым, вторым и третьим остатками фосфорной кислоты являются макроэргическими (в отличие от ковалентных связей они обозначаются волнистой линией ~). Вещества, несущие в своем составе макроэргические связи – макроэргические соединения.
При гидролизе АФТ высвобождается энергия. Гидролиз АТФ возможен двумя путями:
АТФ + Н2 О АДФ + Н3 Р О4 + 32,5 кДж.
АТФ + Н2 О АМФ + Н4 Р2 О7 + 34,7 кДж.
Анаболизм и катаболизм — Студопедия
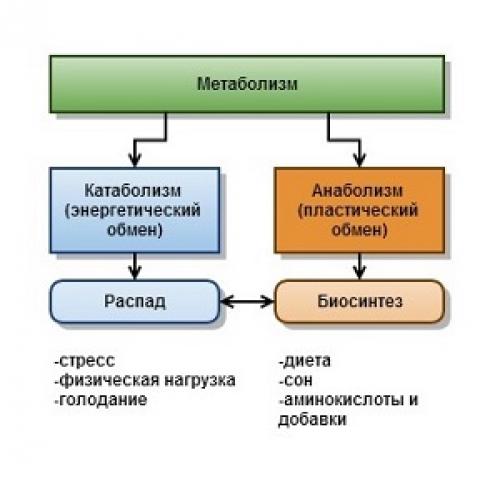
Основными метаболическими процессами являются анаболизм (ассимиляция) и катаболизм (диссимиляция).
Анаболизм, или ассимиляция (от лат. assimilatio — уподобление), представляет собой эндотермический процесс уподобления поступающих в клетку веществ веществам самой клетки. Она является «созидательным» метаболизмом.
Важнейшим моментом ассимиляции является синтез белков и нуклеиновых кислот. Частным случаем анаболизма является фотосинтез, который представляет собой биологический процесс, при котором органическое вещество синтезируется из воды, двуокиси углерода и неорганических солей под влиянием лучистой энергии Солнца. Фотосинтез в зеленых растениях является автотрофным типом обмена.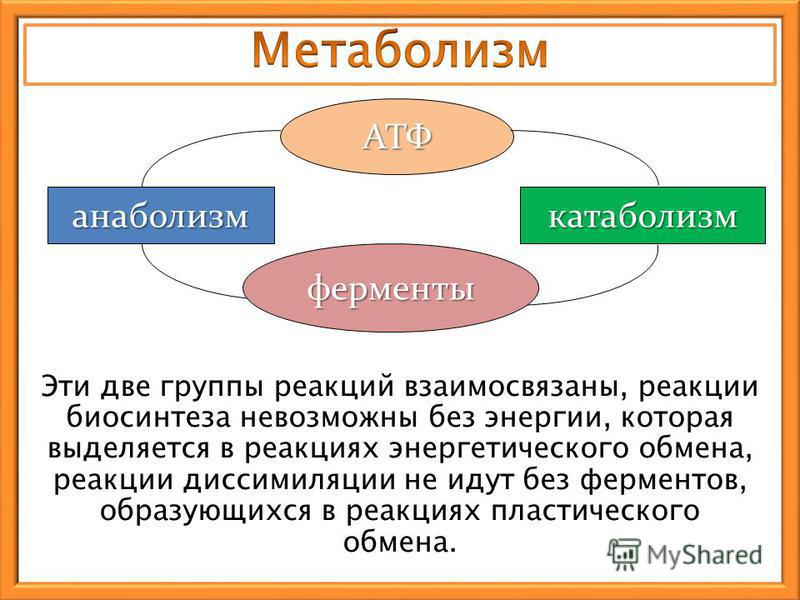
Катаболизм, или диссимиляция (от лат. dissimilis — расподобление), является экзотермическим процессом, при котором происходит распад веществ с освобождением энергии. Этот распад происходит в результате переваривания и дыхания. Переваривание представляет собой процесс распада крупных молекул на более мелкие молекулы, тогда как дыхание является процессом окислительного катаболизма простых Сахаров, глицерина, жирных кислот и дезаминированных аминокислот, в результате которого происходит освобождение жизненно необходимой химической энергии. Эта энергия используется для пополнения запасов аденозинтри-фосфата (АТФ), который является непосредственным донором (источником) клеточной энергии, универсальной энергетической «валютой» в биологических системах. Пополнение запасов АТФ обеспечивается реакцией фосфата (Ф) с аденозиндифосфатом (АДФ), а именно:
АДФ + Ф + энергия ® АТФ
Когда АТФ разлагается на АДФ и фосфат, энергия клетки освобождается и используется для работы в клетке. АТФ представляет собой нуклеотид, состоящий из остатков аденина, рибозы и трифосфата (трифосфатных групп), тогда как аденозиндифосфат (АДФ) имеет лишь две фосфатные группы. Богатство АТФ энергией определяется тем, что его трифосфатный компонент содержит две фос-фоангидридные связи. Энергия АТФ превышает энергию АДФ на 7000 ккал/моль. Этой энергией обеспечиваются все биосинтетические реакции в клетке в результате гидролиза АТФ до АДФ и неорганического фосфата. Итак, цикл АТФ-АДФ является основным механизмом обмена энергии в живых системах.
АТФ представляет собой нуклеотид, состоящий из остатков аденина, рибозы и трифосфата (трифосфатных групп), тогда как аденозиндифосфат (АДФ) имеет лишь две фосфатные группы. Богатство АТФ энергией определяется тем, что его трифосфатный компонент содержит две фос-фоангидридные связи. Энергия АТФ превышает энергию АДФ на 7000 ккал/моль. Этой энергией обеспечиваются все биосинтетические реакции в клетке в результате гидролиза АТФ до АДФ и неорганического фосфата. Итак, цикл АТФ-АДФ является основным механизмом обмена энергии в живых системах.
К живым системам применимы два закона термодинамики.
В соответствии с первым законом термодинамики (законом сохранения энергии) энергия на протяжении химических и физических процессов не создается, не исчезает, а просто переходит из одной формы в другую, пригодную в той или иной мере для выполнения работы, т. е. использование энергии для выполнения какой-либо работы или переход энергии из одной формы в другую не сопровождается изменением (уменьшением или увеличением) общего количества энергии.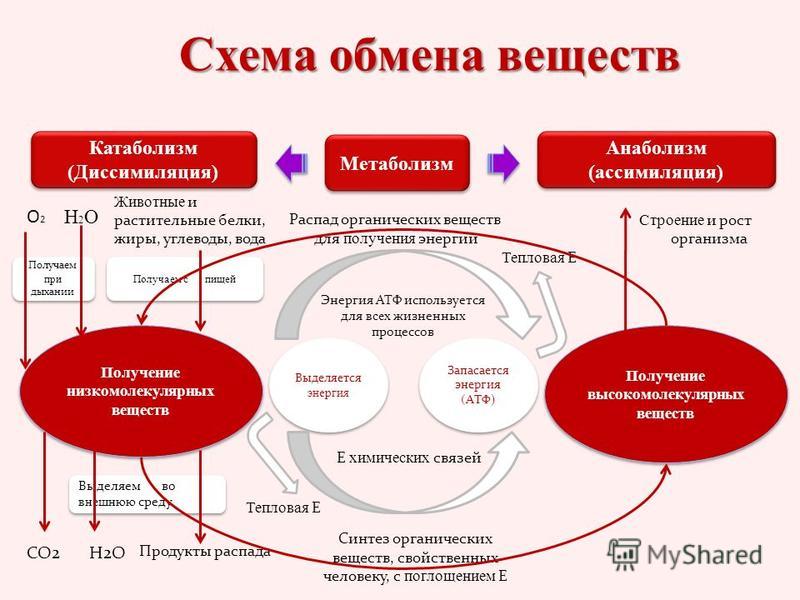 Имея в виду глобальные категории, можно сказать, что вопреки любым физическим или химическим изменениям во Вселенной, количество энергии в ней останется неизменным.
Имея в виду глобальные категории, можно сказать, что вопреки любым физическим или химическим изменениям во Вселенной, количество энергии в ней останется неизменным.
В соответствии со вторым законом термодинамики физические и химические процессы протекают в направлении необратимого перехода полезной энергии в хаотическую, неупорядоченную форму и установления равновесия между упорядоченным состоянием и хаотическим, неупорядоченным. По мере приближения к установлению равновесия между упорядоченностью и неупорядоченностью и к остановке процесса происходит уменьшение свободной энергии, т.е. той порции общей (полезной) энергии, которая способна производить работу при постоянной температуре и постоянном давлении. Когда количество свободной энергии уменьшается, то повышается та часть общей внутренней энергии системы, которая является мерой степени случайности и неупорядоченности (дезорганизации) и называется энтропией. Другими словами, энтропия есть мера необратимого перехода полезной энергии в неупорядоченную форму.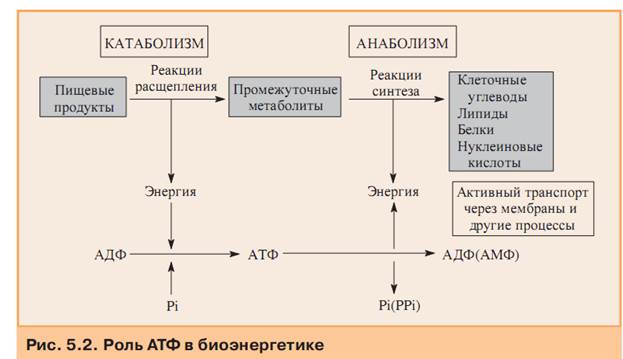 Таким образом, естественная тенденция любой системы направлена на повышение энтропии и уменьшение свободной энергии, которая является самой полезной термодинамической функцией. Живые организмы являются высокоупорядоченными системами. Для них характерно содержание очень большого количества информации, но они бедны энтропией.
Таким образом, естественная тенденция любой системы направлена на повышение энтропии и уменьшение свободной энергии, которая является самой полезной термодинамической функцией. Живые организмы являются высокоупорядоченными системами. Для них характерно содержание очень большого количества информации, но они бедны энтропией.
Если Вселенная представляет собой реакционные системы, под которыми понимают совокупность веществ, благодаря которым протекают физические и химические процессы, с одной стороны, и окружающую среду, с которой реакционные системы обмениваются информацией, с другой стороны, то в соответствии со вторым законом термодинамики в ходе физических процессов или химических реакций энтропия Вселенной увеличивается. Метаболизм живых организмов не сопровождается возрастанием внутренней неупорядоченности, т. е. для живых организмов не характерны возрастные энтропии. В любых условиях все организмы, начиная от бактерий и заканчивая млекопитающими, сохраняют упорядоченный характер своего строения.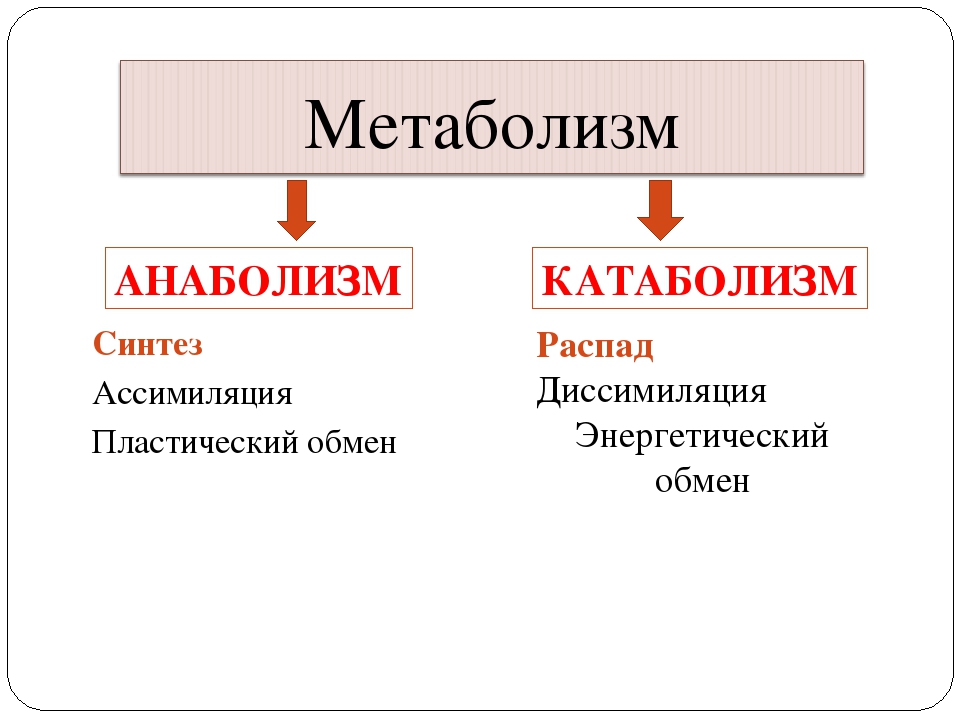 Однако для самой энтропии характерно то, что она возрастает в окружающей среде, причем непрерывное возрастание энтропии в окружающей среде обеспечивается существующими в среде живыми организмами. Например, для извлечения свободной энергии анаэробные организмы используют глюкозу, которую они получают из окружающей среды и окисляют молекулярным кислородом, проникающим тоже из среды. При этом конечные продукты окислительного метаболизма (СО2 и H2O) поступают в среду, что и сопровождается возрастанием энтропии среды, которое частично происходит из-за рассеивания тепла. Возрастание энтропии в этом случае повышается, кроме того за счет возрастания количества молекул после окисления (C6H12O6 + 6O2 ® 6СО2 + 6Н2О), т. е. образование из 7 молекул 12 молекул. Как видно, молекулярная неупорядоченность ведет к энтропии.
Однако для самой энтропии характерно то, что она возрастает в окружающей среде, причем непрерывное возрастание энтропии в окружающей среде обеспечивается существующими в среде живыми организмами. Например, для извлечения свободной энергии анаэробные организмы используют глюкозу, которую они получают из окружающей среды и окисляют молекулярным кислородом, проникающим тоже из среды. При этом конечные продукты окислительного метаболизма (СО2 и H2O) поступают в среду, что и сопровождается возрастанием энтропии среды, которое частично происходит из-за рассеивания тепла. Возрастание энтропии в этом случае повышается, кроме того за счет возрастания количества молекул после окисления (C6H12O6 + 6O2 ® 6СО2 + 6Н2О), т. е. образование из 7 молекул 12 молекул. Как видно, молекулярная неупорядоченность ведет к энтропии.
Для живых существ первичным источником энергии является солнечная радиация, в частности видимый свет, который состоит из электромагнитных волн, встречающихся в виде дискретных единиц, называемых фотонами или квантами света.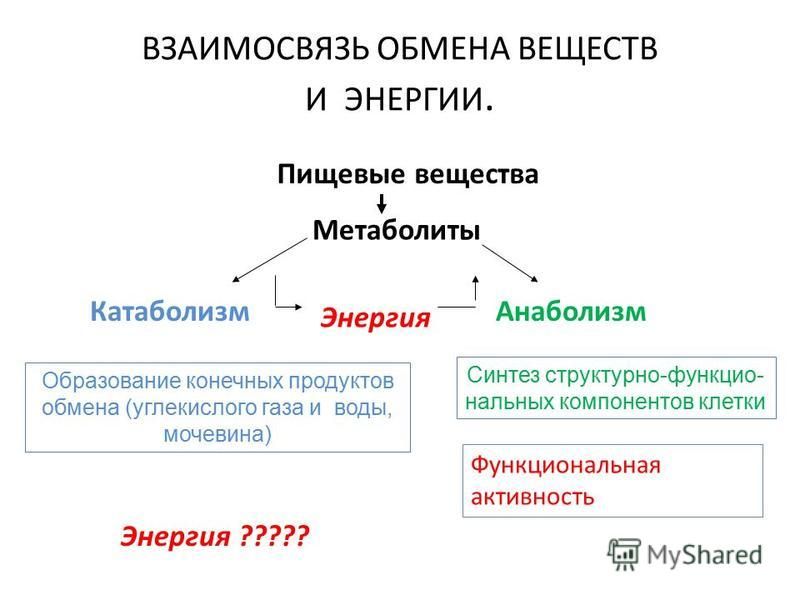 В живом мире одни живые существа способны улавливать световую энергию, другие получают энергию в результате окисления пищевых веществ.
В живом мире одни живые существа способны улавливать световую энергию, другие получают энергию в результате окисления пищевых веществ.
Энергия видимого света улавливается зелеными растениями в процессе фотосинтеза, который осуществляется в хлоропластах их клеток. Благодаря фотосинтезу живые существа создают упорядоченность из неупорядоченности, а световая энергия превращается в химическую энергию, запасаемую в углеводах, являющихся продуктами фотосинтеза. Таким образом, фотосинтезирующие организмы извлекают свободную энергию из солнечного света. В результате этого клетки зеленых растении обладают высоким содержанием свободной энергии.
Получение энергии в результате окисления неорганических веществ происходит при хемосинтезе.
Животные организмы получают энергию, уже запасенную в углеводах, через пищу. Следовательно, они способствуют увеличению энтропии среды. В митохондриях клеток этих организмов энергия, запасенная в углеводах, переводится в форму свободной энергии, подходящей для синтеза молекул других веществ, а также для обеспечения механической, электрической и осмотической работы клеток.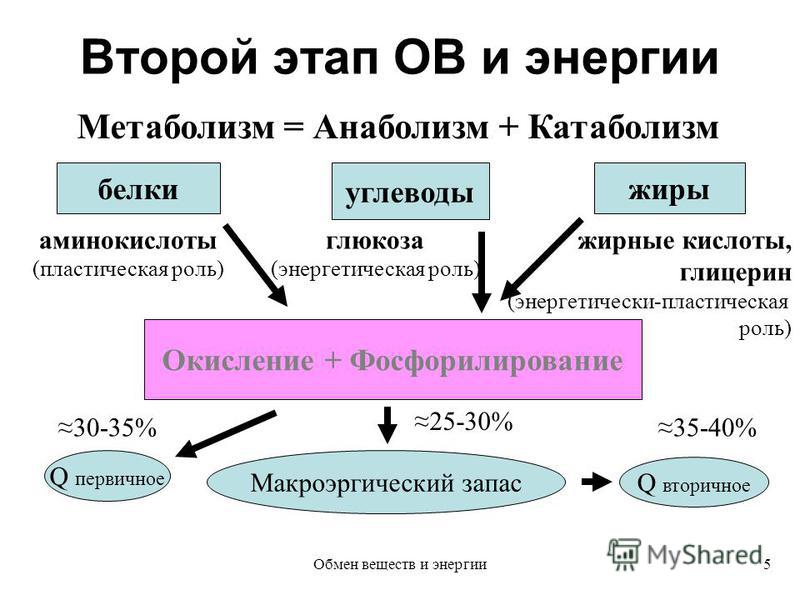 Освобождение энергии, запасенной в углеводах, осуществляется в результате дыхания — аэробного и анаэробного. При аэробном дыхании расщепление молекул, содержащих запасенную энергию, происходит путем гликолиза и в цикле Кребса. При анаэробном дыхании действует только гликолиз. Таким образом, жизнедеятельность клеток животных организмов обеспечивается в основном энергией, источником которой служат реакции окисления-восстановления «топлива» (глюкозы и жирных кислот), в процессе которых происходит перенос электронов от одного соединения (окисление) к другому (восстановление). С окислительно-восстановительными реакциями сопряжено фосфорилирование. Эти реакции протекают как при фотосинтезе, так и дыхании.
Освобождение энергии, запасенной в углеводах, осуществляется в результате дыхания — аэробного и анаэробного. При аэробном дыхании расщепление молекул, содержащих запасенную энергию, происходит путем гликолиза и в цикле Кребса. При анаэробном дыхании действует только гликолиз. Таким образом, жизнедеятельность клеток животных организмов обеспечивается в основном энергией, источником которой служат реакции окисления-восстановления «топлива» (глюкозы и жирных кислот), в процессе которых происходит перенос электронов от одного соединения (окисление) к другому (восстановление). С окислительно-восстановительными реакциями сопряжено фосфорилирование. Эти реакции протекают как при фотосинтезе, так и дыхании.
Организм — открытая саморегулирующая система, она поддерживает и реплицирует себя посредством использования энергии, заключенной в пище, либо генерируемой Солнцем. Непрерывно поглощая энергию и вещества, жизнь не «стремится» к равновесию между упорядоченностью и неупорядоченностью, между высокой молекулярной оранизацией и дезорганизацией. Напротив, для живых существ характерна упорядоченность как в их структуре и функциях, так и в превращении и использовании энергии. Таким образом, сохраняя внутреннюю упорядоченность, но получая свободную энергию с солнечным светом или пищей, живые оранизмы возвращают в среду эквивалентное количество энергии, но в менее полезной форме, в основном в виде тепла, которое, рассеиваясь, уходит во Вселенную.
Процессы обмена веществ и энергии подвержены регуляции, причем существует множество регулирующих механизмов. Главнейшим механизмом регуляции метаболизма является контроль количества ферментов. К числу регулирующих механизмов относят также контроль скорости расщепления субстрата ферментами, а также контроль каталитической активности ферментов. Метаболизм подвержен так называемому обратному аллостерическому контролю, заключающемуся в том, что во многих биосинтетических путях первая реакция может быть ингибирована (подавлена) конечным продуктом. Можно сказать, что такое ингибирование происходит по принципу обратной связи.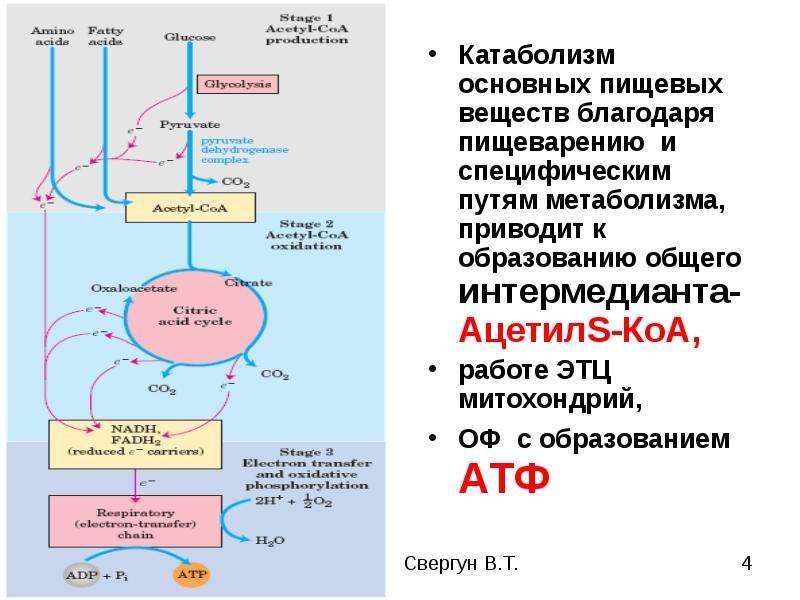 В регуляции обмена веществ и энергии имеет значение и то, что метаболические пути синтеза и распада почти всегда разобщены, причем у эукариотов это разобщение усиливается компартментализацией клеток. Например, местом окисления жирных кислот в клетках являются митохондрии, тогда как их синтез происходит в цитозоле. Многие реакции метаболизма подвержены некоторой регуляции со стороны так называемого энергетического статуса клетки, показателем которого является энергетический заряд, определяемый суммой молярных фракций АТФ и АДФ. Энергетический заряд в клетке всегда постоянен. Синтез АТФ ингибируется высоким зарядом, тогда как использование АТФ стимулируется таким же зарядом.
В регуляции обмена веществ и энергии имеет значение и то, что метаболические пути синтеза и распада почти всегда разобщены, причем у эукариотов это разобщение усиливается компартментализацией клеток. Например, местом окисления жирных кислот в клетках являются митохондрии, тогда как их синтез происходит в цитозоле. Многие реакции метаболизма подвержены некоторой регуляции со стороны так называемого энергетического статуса клетки, показателем которого является энергетический заряд, определяемый суммой молярных фракций АТФ и АДФ. Энергетический заряд в клетке всегда постоянен. Синтез АТФ ингибируется высоким зарядом, тогда как использование АТФ стимулируется таким же зарядом.
Что такое метаболизм? Как анаболизм и катаболизм влияет на массу тела?
Метаболизм — совокупность биохимических процессов, которые протекают в любом живом организме – в том числе в организме человека – и направлены на обеспечение жизнедеятельности. Эти биохимические процессы позволяют нам расти, размножаться, заживлять раны и адаптироваться к меняющимся условиям внешней среды.
Большинство людей использует термин «метаболизм» неправильно, обозначая им либо анаболизм, либо катаболизм.
Слово «метаболизм» происходит от существительного греческого языка «metabole», означающего «перемены», и греческого глагола «metaballein», что в дословном переводе означает «меняться».
Анаболизм и катаболизм
Анаболизмом называют создание материи – последовательность химических реакций, которые строят или синтезируют молекулы из меньших компонентов. Как правило, анаболические реакции сопровождаются потреблением энергии.
Катаболизмом называют разрушение материи – серию реакций химического распада, в ходе которых крупные молекулы расщепляются на меньшие фрагменты. Как правило, процесс протекает с выделением энергии.
Анаболизм
Анаболизм создает материю и потребляет энергию, синтезируя крупные субстанции из небольших компонентов с поглощением энергии в ходе биохимических процессов. Анаболизм, или биосинтез, позволяет организму создавать новые клетки и поддерживать гомеостаз всех тканей.
Организм использует простые молекулы для создания более сложных. Аналогичным образом строитель будет применять простые строительные материалы, например, кирпичи, для возведения здания. Анаболические реакции, протекающие в нашем организме, используют несколько простых веществ и молекул для производства (синтеза) огромного многообразия конечных продуктов. Рост и минерализация костей, набор мышечной массы – примеры анаболизма.
В ходе анаболических процессов из мономеров образуются полимеры. Полимер – это крупная молекула со сложной структурой, состоящая из множества миниатюрных молекул, похожих друг на друга. Эти небольшие молекулы и называют мономерами. Например: аминокислоты, которые являются простыми молекулами (мономерами) в ходе серии анаболических химических реакций образуют протеины, которые являются крупными молекулами со сложной трехмерной структурой (полимер).
К основным анаболическим гормонам относятся:
- Гормон роста – гормон, синтезируемый в гипофизе.
 Гормон роста стимулирует секрецию клетками печени гормона соматомедина, который приводит в действие процессы роста.
Гормон роста стимулирует секрецию клетками печени гормона соматомедина, который приводит в действие процессы роста. - IGF-1 и другие инсулиноподобные факторы роста – гормоны, которые стимулируют образование белка и сульфатов. IGF-1 и IGF-2 участвуют в росте матки и плаценты, а также в начальных стадиях роста плода во время беременности.
- Инсулин – гормон, синтезируемый β-клетками поджелудочной железы. Он регулирует уровень глюкозы в крови. Клетки не могут утилизировать глюкозу без инсулина.
- Тестостерон – мужской гормон, который образуется, главным образом, в яичках. Тестостерон определяет развитие вторичных мужских половых признаков, в частности, низкого голоса и бороды. Также он способствует росту мускулатуры и костной массы.
- Эстроген – женский гормон, который образуется преимущественно в яичниках. Он тоже участвует в укреплении костной ткани и влияет на развитие женских половых признаков, например, молочных желез. Кроме того, эстроген участвует в утолщении внутренней оболочки матки (эндометрий) и других аспектах регуляции менструального цикла.

Катаболизм
Катаболизм разрушает материю и дает нам энергию. В ходе катаболизма крупные молекулярные комплексы распадаются на небольшие молекулы, и этот процесс сопровождается выделением энергии. Катаболизм обеспечивает наше тело энергией, которая необходима для любой физической активности – от клеточного уровня до движений всего тела.
Катаболические химические реакции в живых клетках разрушают крупные полимеры до простых мономеров, из которых они формируются. Например:
- Полисахариды распадаются на моносахариды. Сложные углеводы, такие как крахмал, гликоген и целлюлоза – это полисахариды. Простые углеводы, в частности, глюкоза, рибоза и фруктоза – это моносахариды.
- Нуклеиновые кислоты распадаются на нуклеотиды. Нуклеиновые кислоты являются химической основой жизни и наследственности. В них закодирована вся наша генетическая информация; они служат носителями генетической информации. Примеры – РНК (рибонуклеиновая кислота) и ДНК (дезоксирибонуклеиновая кислота).
 Нуклеиновые кислоты распадаются до пуринов, пиримидинов и пентозы, которая помимо других функций участвует в снабжении нашего организма энергией.
Нуклеиновые кислоты распадаются до пуринов, пиримидинов и пентозы, которая помимо других функций участвует в снабжении нашего организма энергией. - Протеины распадаются до аминокислот. Аминокислоты, образовавшиеся в ходе катаболизма, могут использоваться повторно в анаболических реакциях, идти на синтез других аминокислот или превращаться в другие химические соединения. Иногда белковые молекулы распадаются на аминокислоты для синтеза глюкозы, которая поступает в кровь.
Когда мы едим, наш организм разрушает органические соединения. Этот процесс распада сопровождается выделением энергии, которая в организме хранится в химических связях молекул аденозинтрифосфата (АТФ).
К основным катаболическим гормонам относятся:
- Кортизол – известен также как «гормон стресса», поскольку он участвует в ответной реакции на стресс и тревожность. Гормон продуцируется корой надпочечников, которая является частью надпочечниковой железы. Кортизол повышает артериальное давление и сахар крови, а также подавляет иммунный ответ.
- Глюкагон – гормон, образующийся в α-клетках поджелудочной железы. Он стимулирует распад гликогена в печени, что ведет к повышению уровня сахара крови. Гликоген – углевод, который запасается в печени и используется в качестве топлива во время физической активности. Когда глюкагон выделяется в кровь, он вынуждает клетки печени разрушать гликоген, и тот поступает в кровоток в виде готового топлива (сахар).
- Адреналин – гормон, который образуется в мозговом веществе надпочечниковой железы; адреналин также известен как эпинефрин. Адреналин ускоряет сердечный ритм, увеличивает силу сокращений сердечной мышцы и расширяет бронхиолы в легких. Этот гормон – часть реакции «бей или беги», которая в организме людей и животных является ответом на испуг.
- Цитокины – эти гормоны представляют собой небольшие протеиновые молекулы, которые оказывают специфическое влияние на то, как клетки взаимодействуют между собой, как обмениваются информацией и как себя ведут. Примеры – интерлейкины и лимфокины, которые выделяются при формировании иммунного ответа.

Энергия, хранящаяся в АТФ – это топливо для анаболических реакций. Катаболизм генерирует энергию, которую анаболизм использует для синтеза гормонов, ферментов, сахаров и других субстанций, необходимых для клеточного роста, воспроизведения и регенерации тканей.
Если катаболизм продуцирует больше энергии, чем требуется анаболизму, образуется избыток энергии. Человеческий организм запасает этот избыток энергии в виде жира или гликогена.
Жировая ткань – относительно неактивна по сравнению с мышцами, тканями внутренних органов и другими системами нашего организма. Из-за сравнительно низкой активности жировые клетки для обеспечения жизнедеятельности используют крайне мало энергии в сравнении с другими типами клеток.
Метаболизм и масса тела
Говоря простым языком, масса нашего тела равна результату «катаболизм минус анаболизм». Другими словами, количество энергии, образующейся в нашем организме (катаболизм) минус количество энергии, которую наш организм потребляет (анаболизм).
Избыток энергии аккумулируется в виде жира или гликогена (в виде углеводов энергия хранится преимущественно в печени и мышечной ткани).
При расщеплении одного грамма жира выделяется 9 ккал, а при расщеплении протеина или углеводов – 4 ккал.
Хотя лишний вес чаще всего является результатом накопления организмом энергии в виде жира по причине ее избытка, иногда на метаболизм влияют гормональные нарушения или лежащие в их основе хронические заболевания.
Бытует мнение, что худые люди отличаются «ускоренным обменом веществ», в то время как люди с лишним весом или ожирением страдают от «медленного метаболизма». На самом деле, хронические заболевания, такие как гипотиреоз (низкая активность щитовидной железы), не являются главной причиной ожирения. По данным Государственной Службы Здравоохранения Великобритании, набор веса связан, главным образом, с энергетическим дисбалансом.
Если вы страдаете от лишнего веса или ожирения, целесообразно пройти медицинское обследование и удостовериться в том, что набор массы не вызван эндокринной или соматической патологией.
Кардинально изменить уровень основного обмена – интенсивность метаболизма в состоянии покоя – мы не в силах. Долгосрочные стратегии, такие как набор мышечной массы, могут в конечном итоге дать желаемый результат. Однако определение энергетических потребностей организма с последующей модификацией образа жизни в соответствии с этими потребностями поможет вам намного быстрее уменьшить массу тела.
Энергетические потребности
Масса тела и его композиционный состав. Чем выше масса тела, тем выше потребность в калориях. Верно и то, что люди с высоким соотношением мышц к жировой ткани нуждаются в калориях сильнее, чем лица с аналогичной общей массой, но с меньшим процентом мышечной ткани. Лица с высоким мышечно-жировым соотношением отличаются более высоким уровнем основного обмена, чем люди с аналогичной общей массой, но с меньшим мышечно-жировым соотношением.
Возраст. Когда мы становимся старше, мы сталкиваемся с факторами, которые приводят к сокращению энергетических потребностей. Наша мышечная масса снижается, что приводит к уменьшению мышечно-жирового соотношения. Наш метаболизм постепенно перестраивается, что также влечет за собой уменьшение потребности в калориях.
Наша мышечная масса снижается, что приводит к уменьшению мышечно-жирового соотношения. Наш метаболизм постепенно перестраивается, что также влечет за собой уменьшение потребности в калориях.
Перечисленные ниже возрастные факторы сокращают наши энергетические потребности:
- Гормоны – с возрастом в организме мужчин и женщин образуется меньше тестостерона и эстрогена. Оба гормона участвуют в анаболических процессах, которые потребляют энергию. Синтез человеческого гормона роста, который оказывает колоссальное влияние на анаболические реакции, также уменьшается с возрастом. Когда мы стареем, баланс смещается от анаболических гормонов в сторону катаболических, что резко повышает предрасположенность к набору веса, причем за счет жировой ткани, а не мышц.
- Менопауза – когда женщины приближаются к периоду менопаузы, падает выработка гормонов, которые заставляют организм сжигать больше энергии. Большинство женщин обнаруживает, что сбросить вес в этот период очень проблематично.
 Впрочем, эксперты считают, что менопаузальный и постменопаузальный набор веса лишь отчасти вызван гормональными изменениями. Другие возрастные факторы, в частности, снижение физической активности и несбалансированное питание, оказывают на массу тела гораздо большее влияние.
Впрочем, эксперты считают, что менопаузальный и постменопаузальный набор веса лишь отчасти вызван гормональными изменениями. Другие возрастные факторы, в частности, снижение физической активности и несбалансированное питание, оказывают на массу тела гораздо большее влияние. - Физическая активность – с возрастом люди, как правило, не так активны, как были в молодости. Объясняется это не только более размеренным образом жизни. Большинство людей, которые в молодости зарабатывали тяжелым физическим трудом, после 45 переходят на сидячую работу. Это может быть связано с продвижением по службе, которое имеет место во многих отраслях, например, в армии, полиции, пожарной службе, а также переподготовкой, переходом на принципиально иную работу или ранним выходом на пенсию.
- Теория накопления отходов жизнедеятельности – когда мы становимся старше, растет число клеток с конечными продуктами жизнедеятельности, что, по всей видимости, негативно сказывается на интенсивности обменных процессов.

Пол. Мужчины обладают более высоким уровнем основного обмена, чем женщины, что объясняется большим процентом мышечной ткани в мужском организме. Это значит, что среднестатистический мужчина сжигает больше калорий, чем среднестатистическая женщина его возраста с аналогичной массой тела.
Как сбросить вес?
Сначала вам следует определить суточную потребность организма в калориях и убедиться в отсутствии хронических заболеваний, которые могут быть причиной прибавки массы тела. После этого вы должны сфокусироваться на трех ключевых факторах, влияющих на потерю веса и последующую стабилизацию идеальной массы вашего тела. Те же факторы влияют и на обмен веществ – это физическая активность, диета (рацион питания) и сон.
Значение сна
Если вы не высыпаетесь, нарушается нейроэндокринный контроль чувства голода и насыщения. Следствием этого становится переедание и снижение чувствительности тканей к инсулину, что в свою очередь повышает риск развития диабета 2 типа.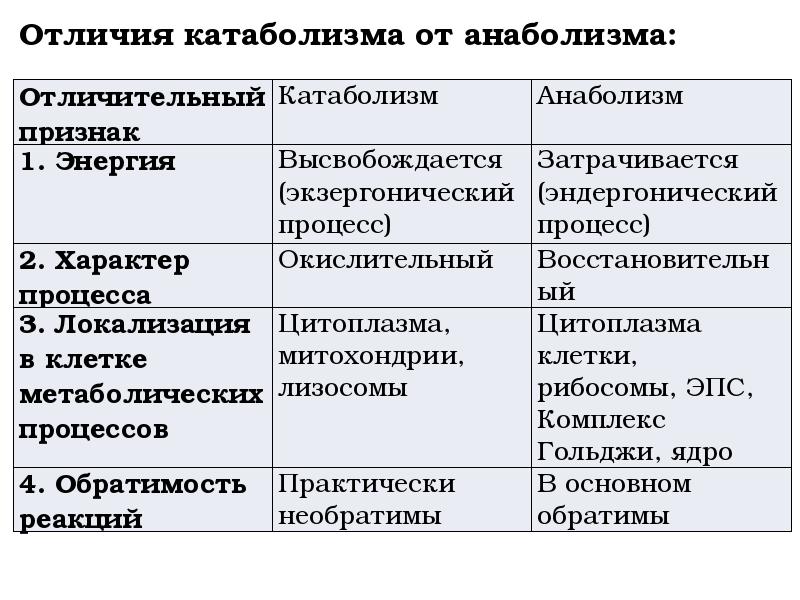 Любой из этих факторов ведет к набору массы тела.
Любой из этих факторов ведет к набору массы тела.
Многочисленные клинические исследования показали, что лишение человека сна нарушает способность организма регулировать пищевое поведение (аппетит) из-за снижения концентрации лептина – гормона, который сообщает нам, что мы съели достаточно.
Ученые, участвовавшие в Интегративном Проекте Здоровья Сердца в Военном Медицинском Центре в Уолтер Рид, пришли к выводу, что существует прямая связь между индексом массы тела (ИМТ) и продолжительностью и качеством сна.
«Когда мы проанализировали имеющиеся данные, разделив участников на «любителей поспать» и «страдающих бессонницей», мы обнаружили, что дефициту сна соответствует более высокий ИМТ – 28,3 кг/м2. Для сравнения ИМТ «любителя поспать» в среднем составлял 24,5 кг/м2. Бессонница также снижала эффективность сна, что проявлялось значительными трудностями при засыпании и частыми пробуждениями», — говорит ведущий исследователь, доктор медицины Арн Элиассон.
Ученые из Университета Бристоля (Англия) пришли к выводу, что если ребенок мало спит, у него повышается риск развития ожирения.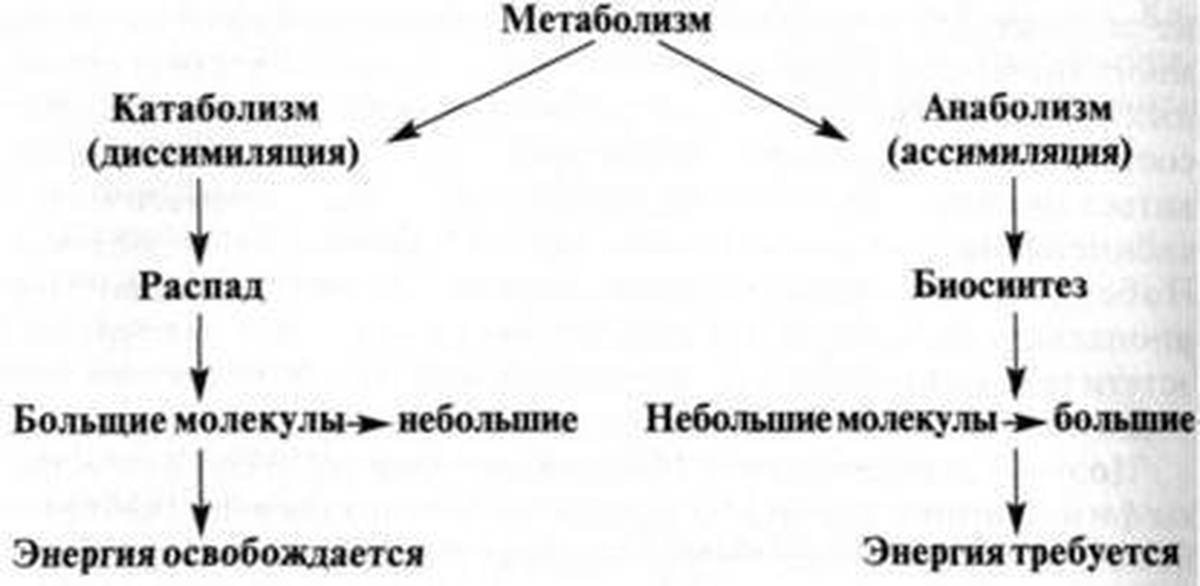 Они считают, что дефицит сна может вести к гормональному дисбалансу, из-за которого дети потребляют больше пищи и в целом питаются неправильно.
Они считают, что дефицит сна может вести к гормональному дисбалансу, из-за которого дети потребляют больше пищи и в целом питаются неправильно.
Исследования также показали, что в организме людей, которые слишком мало спят, повышен уровень грелина. Грелин – это гормон, который синтезируется в желудке и сообщает головному мозгу, что вы голодны.
Сотрудники Колумбийского Университета (Нью-Йорк) пришли к заключению, что недосыпание ведет к снижению толерантности к глюкозе и уменьшению чувствительности к инсулину за счет роста активности симпатической нервной системы, подъема уровня кортизола и уменьшения потребления глюкозы головным мозгом.
Все это резко повышает вероятность набора лишнего веса, а также развития сахарного диабета 2 типа. Те же ученые обнаружили, что у людей, которые спят слишком много (девять часов и более), риск развития диабета тоже повышен.
Ваши шансы набрать лишний вес повышают не только гормональные факторы, связанные с дефицитом сна. Из-за недосыпания вы вряд ли захотите заниматься физкультурой и спортом. Многочисленные эксперименты показали, что люди, которые мало спят, реже придерживаются какой-либо тренировочной программы, и объясняется это тем, что они сильно устают.
Многочисленные эксперименты показали, что люди, которые мало спят, реже придерживаются какой-либо тренировочной программы, и объясняется это тем, что они сильно устают.
Попробуйте следующие меры, которые могут принести вам хороший, полноценный сон:
- Ложитесь спать в одно и то же время.
- Наполните вечерние часы отдыхом и расслаблением.
- Ваша спальня должна быть тихой, темной и немного прохладной.
- Старайтесь получать 7-8 часов непрерывного сна каждую ночь.
- Избегайте продуктов и напитков, которые содержат кофеин.
- Не принимайте обильную пищу непосредственно перед сном. Но и не ложитесь в постель голодным.
- Не делайте энергичные упражнения в пределах 4 часов до отхода ко сну (некоторые эксперты говорят о шести часах).
- В выходные дни продолжайте ложиться спать и просыпаться по установленному графику.
Увеличивайте физическую активность
Шестимесячное исследование, проведенное сотрудниками Медицинского Центра Университета Дьюка, изучало влияние тренировок на организмы 53 участников, которые вели малоподвижный образ жизни.
Ученые сфокусировались на 17 биологических показателях, достоверно повышающих риск кардиоваскулярной патологии. Они оценивали объем талии, физическую подготовку, индекс массы тела, уровень холестерина, чувствительность к инсулину и индикаторы метаболического синдрома – предшественника сахарного диабета 2 типа.
В эксперименте оценивалось три уровня физической активности: эквивалент 20 км ходьбы в неделю, 20 км легкой пробежки и 30 км легкой пробежки в неделю. Участники занимались на беговой дорожке, эллипсоидном тренажере или велоэргометрах под наблюдением исследователей.
Ученые не только обнаружили значительные улучшения к концу исследования, но и пришли к выводу, что интенсивность нагрузки не является решающим фактором.
Вот что говорит руководитель исследования, доктор Дженнифер Роббинс: «Если смотреть на группу в целом, мы обнаружили, что общий положительный эффект был достигнут не только в группе с максимальной интенсивностью нагрузки. Людей должен приободрять тот факт, что они не обязаны выдерживать высокоинтенсивные тренировки, чтобы получать пользу от физических упражнений».
Все упражнения можно разделить на три большие категории
Аэробное упражнение
Целью аэробного упражнения является улучшение потребления кислорода организмом. Термин «аэробный» тесно связан с кислородом. Определение аэробный применяется к метаболическим процессам, в ходе которых используется кислород (катаболические процессы).
Большинство аэробных упражнений выполняется со средним уровнем интенсивности в течение длительного периода, в отличие от других категорий упражнений. Аэробная тренировка включает разминку, выполнение основных упражнений в течение не менее 20 минут и финальную заминку. В аэробных упражнениях участвуют, главным образом, большие мышечные группы.
20-минутный бег – аэробное упражнение, а вот спринт на 200 метров – нет. Получасовая игра в бадминтон – аэробная активность, при условии, что движения игроков сравнительно непрерывны. Гольф, с другой стороны, не считается аэробной нагрузкой, поскольку нет постоянного учащения сердцебиения в течение продолжительного периода времени.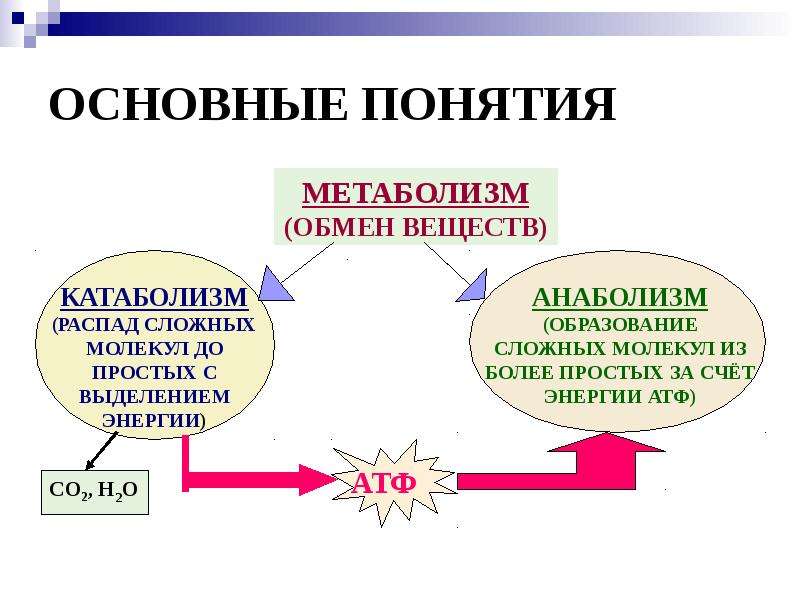
Анаэробное упражнение
Целью анаэробного упражнения является развитие силы, мощности и мускулатуры. Мышцы тренируются с высокой интенсивностью в течение короткого отрезка времени. Под коротким отрезком обычно подразумевается не более двух минут.
Термин анаэробный означает «без воздуха». Анаэробные упражнения увеличивают мышечную силу и нашу способность двигаться с резким ускорением. Вы можете представить анаэробные упражнения как короткие и быстрые, или короткие и интенсивные. Анаэробные упражнения включают силовой тренинг, спринт, быстрые и интенсивные прыжки через скакалку и любые другие быстрые последовательности интенсивных движений.
Поскольку в анаэробных упражнениях кислород не используется для генерации энергии, образуется побочный продукт – молочная кислота. Молочная кислота вызывает мышечную усталость, а потому она должна быть выведена во время восстановления, до того как мышца будет подвергнута очередной анаэробной сессии. Во время восстановительного периода кислород используется для «перезагрузки» мышцы – восполнения внутримышечных запасов энергии, которые были израсходованы во время интенсивного упражнения.
Упражнения на развитие координации и равновесия
Упражнения на развитие координации развивают умение человека резко ускоряться и замедляться, менять направление движения и при этом сохранять равновесие. В теннисе, например, упражнения на развитие координации помогают игроку контролировать свою позицию на корте за счет быстрого возвращения после каждого удара.
Ключевой навык в теннисе – умение занимать правильную позицию на корте, из которой вы можете ударить по мячу максимально эффективно. Хорошая координация не только позволяет теннисисту быстрее подойти к мячу и занять оптимальную для удара позицию, но также помогает лучше сгруппироваться в момент удара по мячу.
Вы должны комбинировать два типа упражнений
Чтобы извлечь из тренировок максимум пользы, вы должны комбинировать аэробные и анаэробные упражнения. И вы должны заниматься пять раз в неделю.
Исследователи из Университета Хериот-Уотт в Эдинбурге (Шотландия) пришли к выводу, что даже непродолжительная, но регулярная и интенсивная нагрузка, например, короткая сессия из четырех-шести 30-секундных высокоинтенсивных спринтов на велотренажере раз в два дня, значительно улучшает способность организма утилизировать сахара.
Диета и питание
Учет калорийности рациона
Ежедневный учет количества потребляемых вами калорий очень важен для контроля массы тела, особенно если вы хотите сбросить вес.
Доказано, что резкое ограничение калорийности рациона неэффективно в долгосрочной перспективе. Экстремальное сокращение калорийности пищи может вынудить организм перестроить метаболизм так, что расходоваться будет намного меньше энергии, а любой источник энергии будет моментально запасаться в жировой ткани. Низкокалорийные диеты часто негативно сказываются на мотивации, что приводит к перееданию после выхода из диеты.
Если только ваша экстремально низкокалорийная диета не разработана квалифицированным диетологом, нутриционистом или профессиональным врачом, велик риск истощения, которое не только вредит здоровью, но и меняет ход обменных процессов таким образом, что достигнуть поставленной цели вам будет еще сложнее.
В США и Великобритании самый высокий процент людей, которые обращаются к этим разрушительным диетам.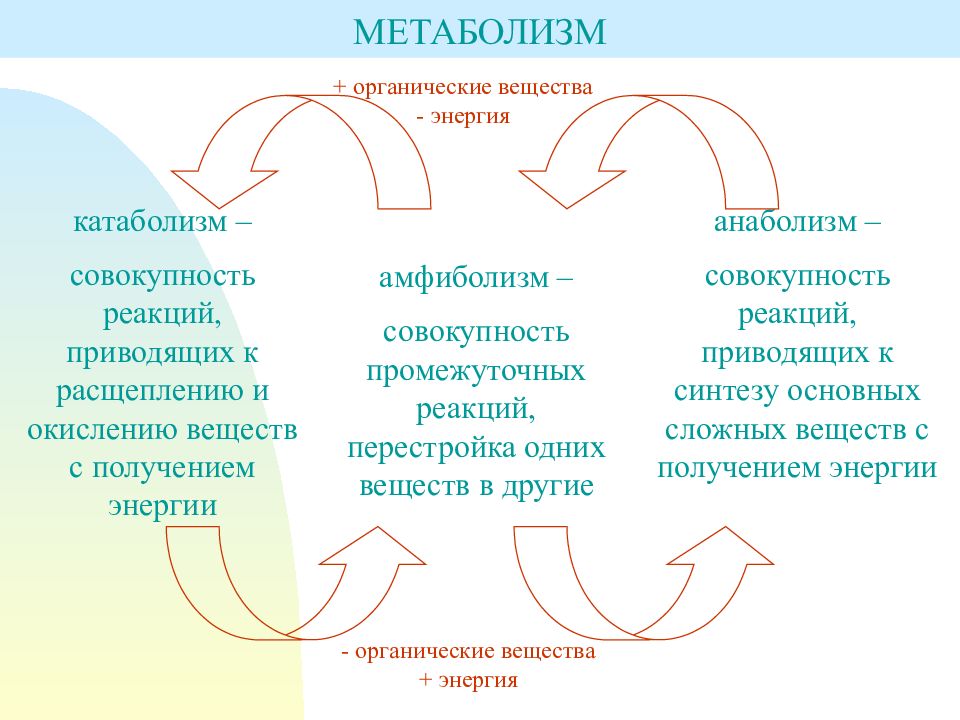 Если бы они были эффективны, Штаты не были бы мировым лидером по числу лиц с ожирением, а Великобритания не лидировала бы по этому показателю в Европе. Из тех, кто сидел на экстремальных диетах, львиная доля по-прежнему страдает ожирением, и лишь немногим удалось вернуться к нормальной массе тела.
Если бы они были эффективны, Штаты не были бы мировым лидером по числу лиц с ожирением, а Великобритания не лидировала бы по этому показателю в Европе. Из тех, кто сидел на экстремальных диетах, львиная доля по-прежнему страдает ожирением, и лишь немногим удалось вернуться к нормальной массе тела.
Здоровая диета
Здоровая диета – хорошо сбалансированная диета. Она должна включать:
Продукты из цельного зерна. Цельное зерно, в отличие от хлопьев, все еще содержит отруби и зародыш в первозданном виде. Продукты из цельного зерна богаты клетчаткой, минералами и витаминами. В процессе переработки зерна отруби и зародыш удаляются из продукта.
Цельнозерновые продукты, включая хлеб, макаронные изделия и крупы, должны производиться из 100% цельного зерна. К продуктам и муке из цельного зерна относится 100% цельнозерновая пшеница, нешлифованный рис, гречиха, овсяная крупа, спельта и дикий рис.
Фрукты и овощи. Фрукты и овощи содержат очень много витаминов, минеральных элементов и клетчатки – эти нутриенты как воздух нужны вашему организму для нормальной жизнедеятельности.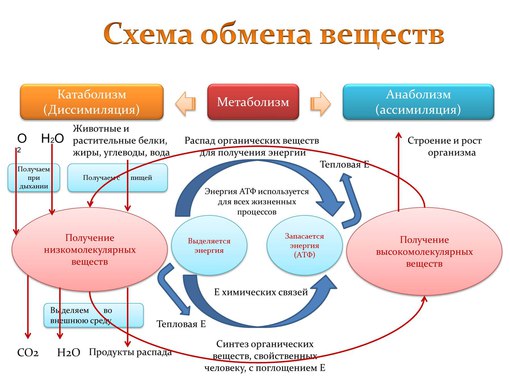 Многочисленные исследования доказали, что богатый фруктами и овощами рацион может защитить от развития болезней сердца, сахарного диабета 2 типа и даже рака.
Многочисленные исследования доказали, что богатый фруктами и овощами рацион может защитить от развития болезней сердца, сахарного диабета 2 типа и даже рака.
Большинство организаций здравоохранения планеты рекомендует нам ежедневно получать пять порций фруктов и овощей. Это могут быть свежие, замороженные, консервированные или высушенные фрукты и овощи. Под порцией следует понимать один большой фрукт, например, яблоко, манго или банан, или три столовых ложки овощей.
Также это может быть стакан 100% фруктового или овощного сока. Заметьте, фруктовый или овощной сок – это одна порция, вне зависимости от ее объема. Бобовые и зернобобовые культуры также можно считать одной порцией.
Протеин. Протеин жизненно необходим для роста и регенерации тканей нашего организма. Богатые протеином продукты содержат еще и незаменимые микроэлементы, например, железо, магний и цинк, плюс витамины группы B. Государственная Служба Здравоохранения Великобритании сообщает, что протеин должен составлять около 20% нашего рациона. Хорошим источником протеина может стать мясо, птица, рыба, яйца, бобы, орехи, кворн (заменитель мяса) и соя (в том числе тофу).
Хорошим источником протеина может стать мясо, птица, рыба, яйца, бобы, орехи, кворн (заменитель мяса) и соя (в том числе тофу).
Диетологи настоятельно рекомендуют сливать масло и срезать жир с мяса после приготовления. С птицы необходимо удалять кожу. Не вегетарианцам нутриционисты советуют есть рыбу не реже двух раз в неделю, выбирая по возможности сорта, богатые омега жирами, например, форель, свежий тунец, сардины, скумбрию и лосось. В процессе консервации из тунца удаляются эссенциальные жиры, а потому только свежий тунец считается жирной рыбой. Рыбу и мясо желательно не жарить, а готовить в микроволновке, на гриле или запекать.
Веганам, которые не едят никаких продуктов животного происхождения, можно получать протеин из орехов, семян, сои, бобов и кворна. Дополнительно веганам стоит принимать пищевые добавки с цинком и витамином B12, так как эти продукты содержат их в недостаточном количестве.
Кальций (молочные или растительные продукты). Молочные продукты считаются хорошим источником кальция, который необходим для здоровья костей и зубов. К молочным продуктам относится молоко, йогурты, сыр и некоторые продукты из соевого молока. Диетологи говорят, что мы должны выбирать нежирные молочные продукты. Люди, которые не едят продукты животного происхождения, могут получить кальций из брокколи, белокочанной капусты, соевого молока и йогурта с добавлением кальция.
К молочным продуктам относится молоко, йогурты, сыр и некоторые продукты из соевого молока. Диетологи говорят, что мы должны выбирать нежирные молочные продукты. Люди, которые не едят продукты животного происхождения, могут получить кальций из брокколи, белокочанной капусты, соевого молока и йогурта с добавлением кальция.
Жиры и углеводы. Стремитесь к качественным жирам, таким как оливковое масло, авокадо или рыбий жир. Избегайте насыщенных жиров, которые содержатся в сливках, жареных блюдах и мясе. Также держитесь подальше от транс-жиров – жиров, полученных искусственным путем. Старайтесь не добавлять сахар в свои блюда, избегайте сладких газированных напитков. В нашей пище и так достаточно углеводов.
Читайте также
Что такое анаболизм и катаболизм
Здравствуйте дорогие читатели, сегодня я хотел бы рассказать о таких важный понятиях как анаболизм, катаболизм и метаболизм (обмен веществ). Так как все о них уже слышали, но не все знают, что они означают. Поэтому давайте разберемся, что же это такое.
Поэтому давайте разберемся, что же это такое.
Метаболизм
Это набор химических реакций, которые поддерживают жизнь живого организма (размножение и рост). Метаболизм делится на 2 вида: анаболизм и катаболизм, поэтому одно без другого не может существовать. Что бы было понятнее – рассмотрим метаболизм на примере живого существа (человека, животного итак далее):
В процессе эволюции живые организмы научились выживать за счет того, что у них развился механизм накопления и сжигания внутреннего вещества (анаболизм и катаболизм). Это можно представить в виде агрегата работающего на солнечных батареях. Есть солнце, все крутится и вертится, а лишняя энергия запасается в аккумуляторы (анаболизм). Нет солнца, начинают работать батареи (катаболизм). И если долго не будет солнца, то наш механический прообраз человеческого организма — остановится.
Поэтому жизнь устроена почтитак, если рассматривать ее в первом приближении. Наш организм основан на том же принципе, что даже если после длительного не поступления в организм энергии (пищи), он не выйдет из строя. Живые существа научились частично разрушать себя, используя высвободившуюся энергию для продолжения движения, чтобы найти пищу. Пока ученые не сумели сделать такой механизм в лаборатории и, наверное, они не скоро научатся. Природе для этого понадобилось огромный период времени…
Живые существа научились частично разрушать себя, используя высвободившуюся энергию для продолжения движения, чтобы найти пищу. Пока ученые не сумели сделать такой механизм в лаборатории и, наверное, они не скоро научатся. Природе для этого понадобилось огромный период времени…
Анаболизм и катаболизм
Теперь, когда все примерно стало понятно с метаболизмом, давайте разберемся с терминами анаболизма и катаболизма.
Анаболизм – это процесс создания (синтез) новых веществ, клеток и тканей. Например, создание мышечных волокон, новых клеток, накопление жиров, синтез гормонов и белков.
Катаболизм – это процесс обратный анаболизму, то есть расщепление сложных веществ на более простые, и распад тканей и клеток. Например расщепление (разрушение) жиров, продуктов питания итак далее.
Не надо быть провидцем, чтобы понять, что эти два процесса должны уравновешивать друг друга. Поэтому только тогда живое существо сможет сохранять свое здоровье и жизнь. На этом месте можно было бы сделать паузу и спросить себя, а зачем мне все это нужно знать? Все ведь так хорошо устроено.
Так-то оно так, но есть беспокойные люди, которым очень хочется нарушить это равновесие, чтобы получить, например, увеличение мышечной массы. Они готовы часами доводить себя на тренировках в тренажерных залах, чтобы увеличить свой бицепс или косую мышцу. Даже придуман был для этого особый вид спорта – бодибилдинг. Так вот, если человек занимаясь, немного представляет, что внутри его организма происходит это одно, а когда делает это по незнанию, это другое.
В жизни тоже много ситуаций, которые хочется как-то объяснить, чтобы понять и принять правильное решение. Возьмем простой пример: молодая и стройная девушка, кушает все подряд и при этом не набирает в весе. Прошло пару десятков лет и вдруг все изменилось — она пополнела.
А связано это с тем, что с годами обменные процессы (метаболизм) замедляются, а это и приводит к накоплению лишнего веса, если как следует не следить за собой (правильное питание и подвижный образ жизни). Однако не у всех так происходит, есть счастливчики, которые всю жизнь кушают все подряд, не занимаются спортом и остаются стройными…
Анаболические стероиды
Это гормональные препараты, которые используются спортсменами для увеличения мышечной массы, но данные препараты очень опасны для здоровья. Так как они вмешиваются в анаболический процесс, то есть создание новых клеток и тканей, что ведет к нарушению гормонального фона (гормональной системы). В результате такого вмешательства могут возникнуть проблемы со здоровьем, таких органов как: сердце, печень и почки.
Так как они вмешиваются в анаболический процесс, то есть создание новых клеток и тканей, что ведет к нарушению гормонального фона (гормональной системы). В результате такого вмешательства могут возникнуть проблемы со здоровьем, таких органов как: сердце, печень и почки.
Но так же есть и «катаболические» стероиды, которые применяют в медицине для лечения различных тяжелых заболеваний, однако ими пользуются и спортсмены для ускоренного сжигания жиров (сушка). Онитак же вредны и вмешиваются в гормональную систему, действие таких препаратов-обратно действию (обратно пропорционально) анаболических. Поэтому занимайтесь «чистым» спортом без каких либо препаратов и будите здоровы.
Подведем итог. Метаболизм – это процесс химических реакция который поддерживает жизнь (размножение и рост), а состоит метаболизм из двух составляющих: анаболизм (создание новых веществ и клеток) и катаболизм (расщепления сложных веществ на более простые). И одно без другого не может существовать (анаболизм и катаболизм), так как баланс (равновесие) – это есть жизнь (гармония).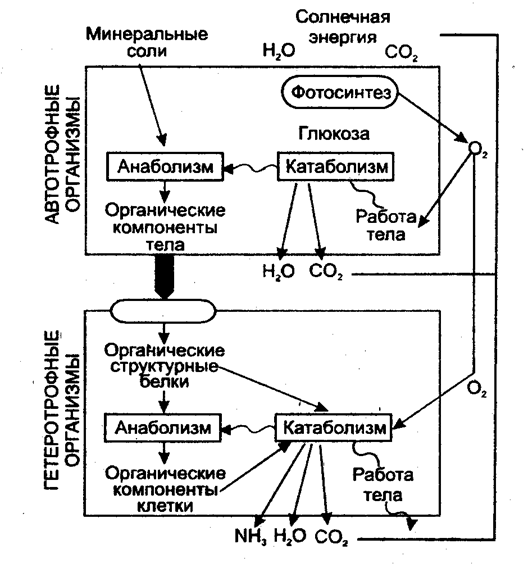 Занимайтесь «чистым» спортом без каких либо анаболических и катаболических препаратов, которые гробят ваше здоровье.
Занимайтесь «чистым» спортом без каких либо анаболических и катаболических препаратов, которые гробят ваше здоровье.
Занимайтесь спортом, питайтесь правильно – успехов Вам!
виды и факторы, влияющие на него
Хороший метаболизм – обязательное условие для построения здорового и спортивного тела. О том, что такое анаболизм и катаболизм, и о причинах, по которым Ваш обмен веществ может быть нарушен, читайте в статье.
Каждый раз, когда речь заходит о похудении, говорят о метаболизме и о том, что его нужно увеличить. Полные люди зачастую видят причину своего лишнего веса именно в замедленном обмене веществ. Да и говоря о наборе массы, вопросы скорости метаболизма тоже в числе лидирующих. Так почему же он так важен?
Что такое метаболизм?
Метаболизм (или обмен веществ) – это процесс переработки питательных веществ, поступающих в организм с пищей, в энергию. Процессам метаболизма подвергаются абсолютно все вещества в организме. Это и белки, и жиры, и углеводы и любые микро-, макроэлементы, и минеральные вещества.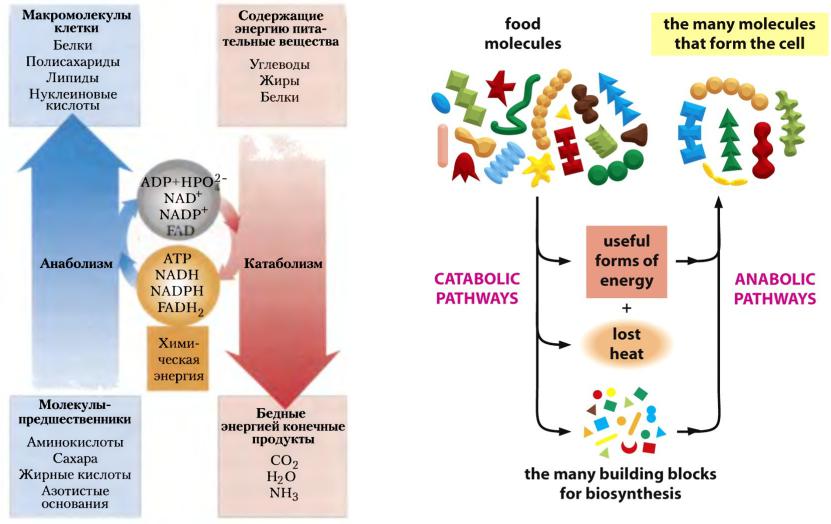 Каждая клетка Вашего тела участвует в обменных процессах.
Каждая клетка Вашего тела участвует в обменных процессах.
Виды метаболизма
Существует два основных вида обмена веществ – анаболизм и катаболизм.
Анаболизм – совокупность химических процессов, направленных на создание новых клеток и тканей в организме. Одним из самых ярких примеров анаболизма является рост мышечной массы.
Катаболизм – процессы разрушения и распада сложных веществ до более простых. В процессе катаболизма, как правило, выделяется энергия, которую мы и используем для обеспечения нормальной жизнедеятельности. Когда мы говорим о жиросжигании, мы тоже говорим о катаболизме, ведь нам нужно чтобы энергия для нашей активности была получена именно за счёт расщепления всех трех видов жиров, существующих в теле человека.
Что такое скорость обмена веществ?
Это основной показатель метаболизма. Скорость обмена веществ – это то количество энергии, которое Ваш организм тратит для обеспечения жизнедеятельности. Обычно эта цифра рассчитывается для одного дня и равна количеству килокалорий, необходимых для покрытия энергозатрат. Чем больше этот показатель, соответственно, тем выше скорость Вашего метаболизма.
Чем больше этот показатель, соответственно, тем выше скорость Вашего метаболизма.
Формулы, по которым можно рассчитать скорость метаболизма я приводила в статье «Норма килокалорий»
Метаболизм и жиросжигание
Скорость обмена веществ напрямую связана с успехом в процессе похудения. Ведь чем больше калорий Вы потратите, чем быстрее сможете избавляться от жира (естественно, при условии дефицита калорийности). И в данном случае, нас интересует именно жировой (липидный) обмен, если мы хотим худеть за счёт жира, а не мышц.
Существуют люди с относительно быстрым обменом веществ, которые как правило, могут кушать достаточно много и при этом оставаться очень «сухими». Но так же есть с люди с замедленным метаболизмом, которые кушают умеренно, но быстро набирают жир.
Но не спешите относить себя к тем или другим. На самом деле, на скорость обмена веществ влияет множество факторов (об этом речь пойдёт ниже), в том числе образ жизни и привычки питания. И зачастую те, кто винит в лишнем весе медленный метаболизм, на самом деле просто едят больше, чем им необходимо и ведут малоподвижный образ жизни.
Метаболизм и набор массы
Многие считают, что скорость обмена веществ важна только для тех, кто хочет похудеть, но это не так. При наборе массы огромное значение имеют обе формы метаболизма. Потому что с одной стороны, чтобы мышцы росли, нужно, чтобы процессы анаболизма протекали интенсивнее. Чем выше анаболизм, тем больше рост массы. С другой стороны, очень Важно замедлить процессы катаболизма, т. е не допустить, чтобы для получения энергии происходило расщепление мышечной ткани. Все эти процессы регулируются соответствующе подобранным режимами питания и тренировок.
Основные факторы, влияющие на скорость обмена веществ:
Пол – базальный метаболизм у мужчин выше, чем у женщин.
Возраст – по мере взросления скорость метаболизма постепенно уменьшается. Пик метаболической активности наблюдается в возрасте 20-ти лет. После этого за каждые 10 лет скорость метаболизма снижается на 2-3%.
Гормоны – все обменные процессы и физиологические функции в организме осуществляются по средствам гормональной регуляции. Соответственно, если у Вас наблюдаются какие-то сбои в гормональной системе, то они могут стать не только причиной замедления обмена веществ, но и провоцировать излишнее образование жира.
Соответственно, если у Вас наблюдаются какие-то сбои в гормональной системе, то они могут стать не только причиной замедления обмена веществ, но и провоцировать излишнее образование жира.
Режим питания – частые диеты и нерегулярные приёмы пищи приводят к снижению скорости метаболизма (более подробно об этом я писала в этом материале).
Вода – вода участвует абсолютно во всех процессах организма. Она участвует в процессах терморегуляции, растворяет питательные вещества, выводит вредные продукты обмена. Вывод очевиден: если воды поступает недостаточно, то и процессы обмена протекают медленнее.
Физическая активность – чем более подвижен Ваш образ жизни, тем быстрее проходят обменные процессы в организме.
Тип телосложения – все мы имеем ту или иную генетическую предрасположенность к определённому типу фигуры. Одни больше склонны к полноте, другие наоборот худощавы. Это фактор заложен в каждом человеке при рождении.
Как видите, метаболизм несёт ответственность за постоянство внутренней среды организма и нормальное протекание всех процессов в нём. Именно поэтому, если Вы серьёзно озабочены качеством своего тела, первое, что нужно сделать – нормализовать скорость метаболизма. О том, как это сделать, читайте в материале «Как разогнать метаболизм до предела?».
Именно поэтому, если Вы серьёзно озабочены качеством своего тела, первое, что нужно сделать – нормализовать скорость метаболизма. О том, как это сделать, читайте в материале «Как разогнать метаболизм до предела?».
Понравилась статья? Скажите «спасибо» автору и поделитесь ей в социальных сетях, нажав на соответствующую иконку в правом нижнем углу.
А чтобы получать больше полезной информации каждый день, подпишитесь на наш instagram.
Что такое катаболизм, анаболизм. Какое отношение они имеют к метаболизму и как происходят? :: SYL.ru
В живом организме постоянного происходят различные процессы для обеспечения его жизнедеятельности. Одним из них является обмен веществ (метаболизм), который преобразует потребляемую пищу в энергию. Именно о метаболизме пойдёт речь в данной статье. Мы рассмотрим сущность обменных процессов, уточним их этапы и ответим на вопрос — что такое катаболизм и анаболизм.
Сущность метаболизма и его роль для живых организмов
Для того чтобы живые организмы могли расти, восстанавливаться и размножаться, природа наделила их крайне важной способностью преобразовывать калории из поступающих извне продуктов питания в ценную и нужную для жизни энергию. Совокупность этих биохимических процессов называется обменом веществ или метаболизмом.
Совокупность этих биохимических процессов называется обменом веществ или метаболизмом.
Метаболизм каждой особи может быть выражен количественным коэффициентом, который определяет скорость преобразования организмом пищи в полезную энергию. Установлено, что на активность обменных процессов влияет ряд факторов:
- Пол. У мужчин коэффициент метаболизма на 10-20% больше, чем у женщин.
- Возраст. В целом, метаболизм (анаболизм, катаболизм) быстрее протекает в первые 25-30 лет жизни, после наблюдается снижение скорости обмена веществ на 3% каждое десятилетие.
- Излишний вес приводит к быстрому освоению питательных веществ, которые откладываются в виде жира в мышечных тканях и печени.
- Физические нагрузки ускоряют метаболизм в следующей прогрессии – на 20% в течение первых 2-3 часов, а в последующие сутки — на 5%.
Метаболистические процессы в живых организмах протекает неизбежно в двух противоположных формах: диссимиляции (катаболизм) и ассимиляции (анаболизм).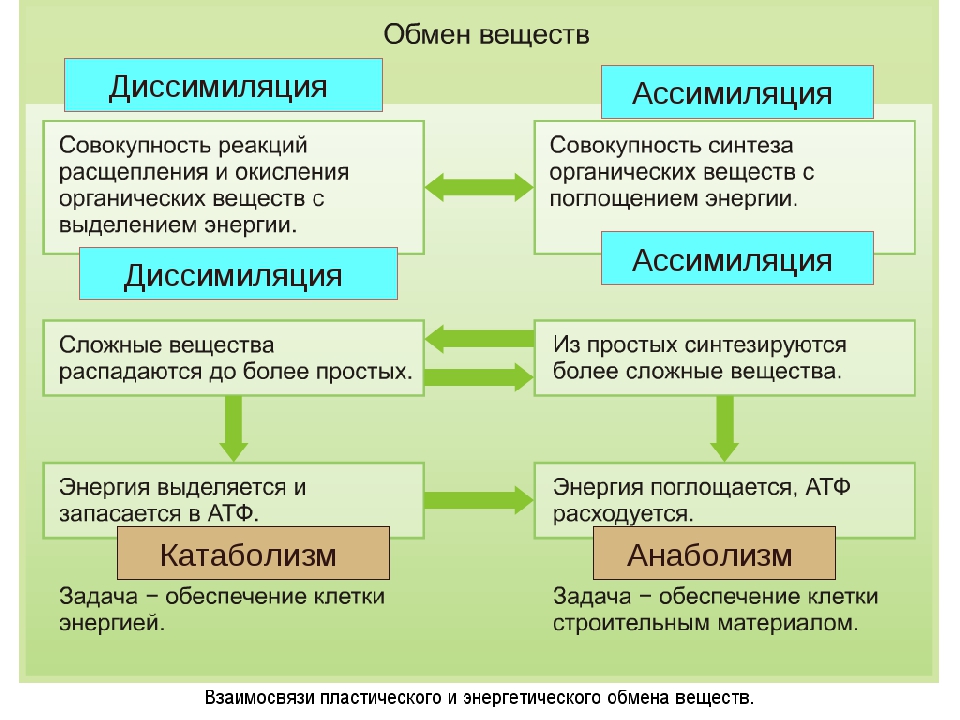 Далее мы рассмотрим эти процессы подробнее.
Далее мы рассмотрим эти процессы подробнее.
Подробнее о том, что такое катаболизм
Катаболизм – это процесс, в ходе которого сложные вещества, в виде совокупности клеток, тканей, органов и прочего, расщепляются на более простые. Процессы катаболизма обязательно сопровождаются образованием и обогащением клеток энергий в виде АТФ, которая впоследствии может быть израсходована на синтез и другие процессы жизнедеятельности, например, движение.
На катаболизм, а точнее на скорость расщепления сложных веществ, влияют следующие гормоны:
- кортизол — вырабатываются в организме вследствие стрессовых ситуаций;
- адреналин, концентрация которого может быть увеличена при голоданиях, а также тогда когда человек испытывает сильные эмоции;
- глюкагон – гормон сжигания жира, активно вырабатывающийся при ограничении количества поступления углеводов из продуктов питания (в диетологии это возможно благодаря диетам с преобладающим потреблением белковой пищи).
Этапы катаболизма
Диссимиляция сложных соединений проходит несколько последовательных этапов, среди которых:
- Расщепление органических молекул жиров, белков и сложных углеводов в мономеры (например, жирные кислоты, аминокислоты, моносахариды).
 Процесс происходит вне клетки – в пищеварительном тракте.
Процесс происходит вне клетки – в пищеварительном тракте. - Попадание мономеров с помощью циркуляции крови и лимфы в тканевые клетки, где процесс расщепления продолжается до образования более мелких структур, таких как ацетильная группа коферментов.
- Окисление под воздействием кислорода в процессе дыхания низкомолекулярных соединений, в результате чего образуется углекислый газ и вода, а также накапливается клеточная энергия АТФ.
Как происходит анаболизм
После того как мы рассмотрели, что такое катаболизм, будет уместно дать понятие и его противоположной форме – анаболизму. Итак, если катаболизм является процессом расщепления полимеров, то анаболизм – это не что иное как синтез простых веществ в сложные соединения, из которых строятся новые клетки и ткани организма.
Анаболизм обеспечивает рост, развитие и регенерацию всех типов тканей организма.
Ассимиляционный метаболизм также проходит в три этапа:
- Вначале низкомолекулярные соединения образуют молекулы-предшественники.

- В следующем этапе эти молекулы преобразовываются в активные формы, при этом затрачивается накопленная при катаболизме клеточная энергия.
- Завершающий этап включает процесс образования строительного материала организма — полимеров, в виде белков, углеводов и жиров.
Взаимосвязь катаболизма и анаболизма
Катаболизм и анаболизм — тесно взаимосвязанные процессы. Первый обеспечивает распад органических соединений до простых веществ и накопление энергии, которое нужно для ассимиляционного обмена. Второй снабжает катаболистические процессы необходимыми ферментами.
Эти две формы обмена непрерывно протекают в живых организмах и могут находиться в двух вариантах взаимодействия:
- в состоянии равновесия;
- в преобладании одного вида над другим.
Сохранение или нарушение равновесия метаболистических процессов зависит от возраста и от психо-эмоционального состояния организма. Так, например, у детей, особенно в первый год жизни, наблюдается преобладание анаболизма над катаболизмом, а у пожилых людей — наоборот.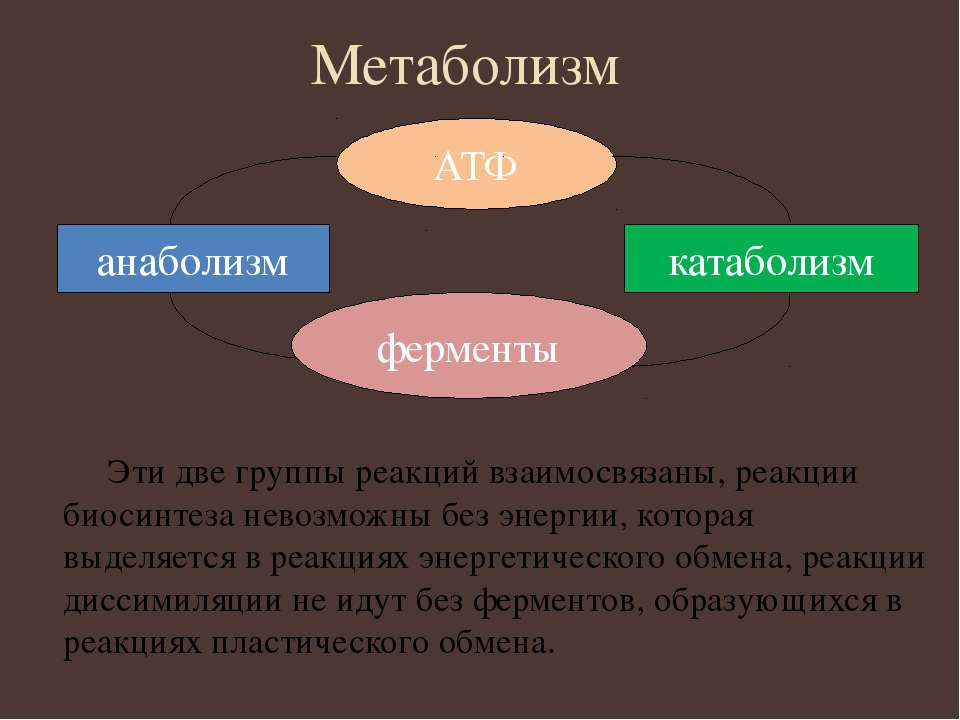
Состояние стресса и физические нагрузки также приводят к смещению равновесия обменных процессов в сторону диссимиляции. Ведь что такое катаболизм? В практическом понимании, это снижение веса и сжигание калорий.
О жизни в метаболическом городе — Strelka Mag
«Метаболизм» человеческих систем стал механизмом обработки их воздействия на окружающую среду. Представляя их в виде фигур химического и энергетического потока, метаболизм позволяет количественно оценить окончательное влияние технологической и социальной деятельности. Но прежде чем действовать, необходимо каким-то образом навести мост между системным и человеческим масштабами.
«Энергетические процессы в промышленном городе подобны процессам в плотном устричном рифе.
— Говард Одум
БЫТЬ МЕТАБОЛИЧЕСКИМ
Афанасий Кирхер, «Вода, текущая под землей». Из Mundus subterraneus, 1665
Быть метаболическим — значит населять ландшафт потоков. В мире, где понимаются как технологическое, так и естественное экологически , метаболизм фактически стал универсальным. Любой вид идентичности, любая ограниченная система в потоке материала может быть определена метаболическим действием.Все потребители и преобразователи материи и энергии — от электростанций до коралловых рифов, от сосен до батарей — можно рассматривать с точки зрения равенства. Метаболизм делает их вычислимыми. Он фиксирует то, что они делают и что они из себя представляют, что в данном конкретном мировоззрении одно и то же.
В мире, где понимаются как технологическое, так и естественное экологически , метаболизм фактически стал универсальным. Любой вид идентичности, любая ограниченная система в потоке материала может быть определена метаболическим действием.Все потребители и преобразователи материи и энергии — от электростанций до коралловых рифов, от сосен до батарей — можно рассматривать с точки зрения равенства. Метаболизм делает их вычислимыми. Он фиксирует то, что они делают и что они из себя представляют, что в данном конкретном мировоззрении одно и то же.
Первоначально биологическая концепция активности текущей жизни — светская замена анима или витальной искры — метаболизм теперь является инструментом. Его применение к продуктам человеческого дизайна — городам, фабрикам, процессам и объектам всех видов — имеет фундаментальное значение для современной оценки воздействия на окружающую среду. Метаболизм сводит свой предмет к взаимосвязи потоков и тем самым позволяет генерировать экологическую валюту. Когда мы говорим о воплощенном углероде той или иной деятельности, мы имеем в виду метаболическое сокращение набора технологических процессов. Когда мы связываем какое-либо социальное или экономическое поведение с балансом воздействия на окружающую среду, мы используем один и тот же прием.
Метаболизм сводит свой предмет к взаимосвязи потоков и тем самым позволяет генерировать экологическую валюту. Когда мы говорим о воплощенном углероде той или иной деятельности, мы имеем в виду метаболическое сокращение набора технологических процессов. Когда мы связываем какое-либо социальное или экономическое поведение с балансом воздействия на окружающую среду, мы используем один и тот же прием.
Учреждение по производству и анализу материальных потоков
Метаболизм — это показатель химического потока.Он описывает потребление и преобразование энергии и химикатов в определенное время и в определенном месте. Метаболизм — это пространственная непрерывность в panta rhei непрерывного движения. Вот и все, что они есть. Стремиться понять город или общество с точки зрения метаболизма, по сути, означает признать, как мало вы еще о нем знаете. Мелкие и сложные внутренние детали не учитываются. Не делается попыток ни пространственной, ни формальной схематизации движущихся частей. Допускаются только входы и выходы; подпись непрекращающейся и непостижимой деятельности по самотворению и поддержанию.
Не делается попыток ни пространственной, ни формальной схематизации движущихся частей. Допускаются только входы и выходы; подпись непрекращающейся и непостижимой деятельности по самотворению и поддержанию.
BANAL — FUTURISTIC
Потребление материалов в мировой экономике, произведено Circle Economy, 2018
Возникнув как радикальное переосмысление городской формы в начале 1960-х годов, метаболизм превратился в простую и неприкрытую загробную жизнь, встроенную в процессы поста. -1990-е гг. Экологический анализ. В рамках таких дисциплин, как Анализ материальных потоков (MFA) и Оценка жизненного цикла (LCA), он конструирует действие и влияние человеческих процессов друг на друга и окружающую среду.Метаболизм является синонимом определения деятельности (будь то работа на алюминиевом заводе в течение года, вождение грузовика по ровной дороге в течение часа или ловля килограмма сардин). Именно потоки между такими системами составляют основную экологическую реальность.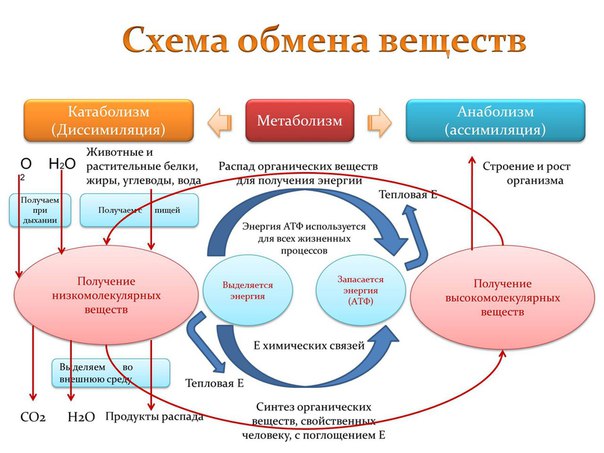 В экологическом прочтении человеческого техно-общества ресурсы, полезные ископаемые, топливо и загрязнители в конечном итоге более значимы, чем предметы, продукты, товары или покупки. Последнее является лишь поверхностным выражением более глубокого движения первого.
В экологическом прочтении человеческого техно-общества ресурсы, полезные ископаемые, топливо и загрязнители в конечном итоге более значимы, чем предметы, продукты, товары или покупки. Последнее является лишь поверхностным выражением более глубокого движения первого.
Как заметила Лидия Каллиполити в «История экологического дизайна », экология как мировоззрение подразумевает эквивалентность технологии и природы. Потоки становятся реальностью как внутри человеческого техно-общества, так и геобиосферы за его пределами. Движение энергии и материала через границу между ними является показателем воздействия человека. Таким образом, потоки принимают температуру атмосферного или экстракционного кризиса. Но источник их диагностической силы — их абстрагирование от человеческой культуры и масштаба — также делает их удручающе нематериальными.Процесс создания видимых потоков, их воссоединения с мизансценой повседневной человеческой реальности — сложный проект.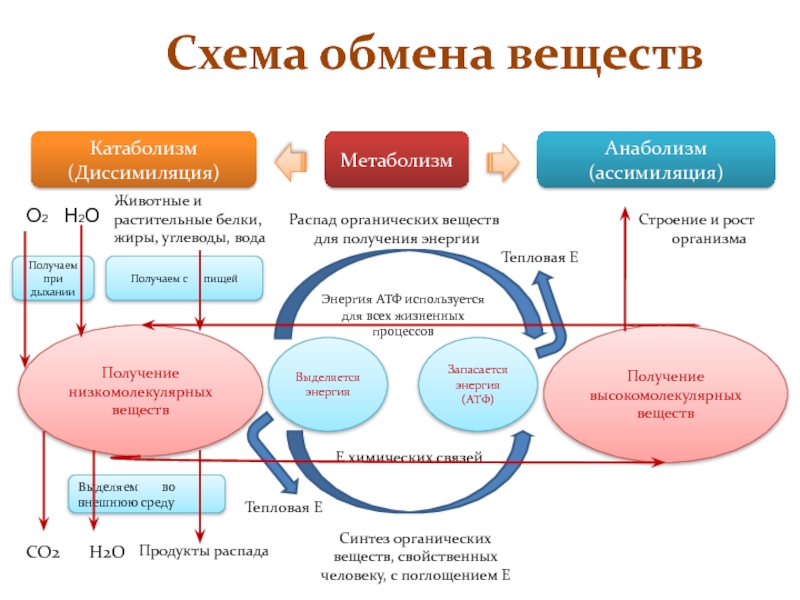 В эпоху, когда дизайн и инвестиции во всем мире все чаще оцениваются по экологическим критериям, это тот период, вокруг которого можно увидеть форму будущего дизайна.
В эпоху, когда дизайн и инвестиции во всем мире все чаще оцениваются по экологическим критериям, это тот период, вокруг которого можно увидеть форму будущего дизайна.
В частности, метаболический город — это поле, в котором этот регистр видимости драматизируется. Для поколения архитекторов 1960-х годов идея городского метаболизма отражала непрерывные изменения в высокотехнологичном городе будущего.То, что их впечатляющий футуризм, рассматриваемый вдоль оси истории, должен соответствовать банальным статистическим процедурам современного анализа окружающей среды, идущим в другую сторону, — это ирония.
Но это несовместимость, которая в значительной степени остается нерешенной и продуктивной. Сфера человеческого социального и технологического метаболизма — это область, в которой интеллект и стратегическая «глупость», форма и бесформенность, материал и медиа — все пересекаются и трансформируются. Для идеи, которая формирует будущее города, особенно недооценивается метаболизм.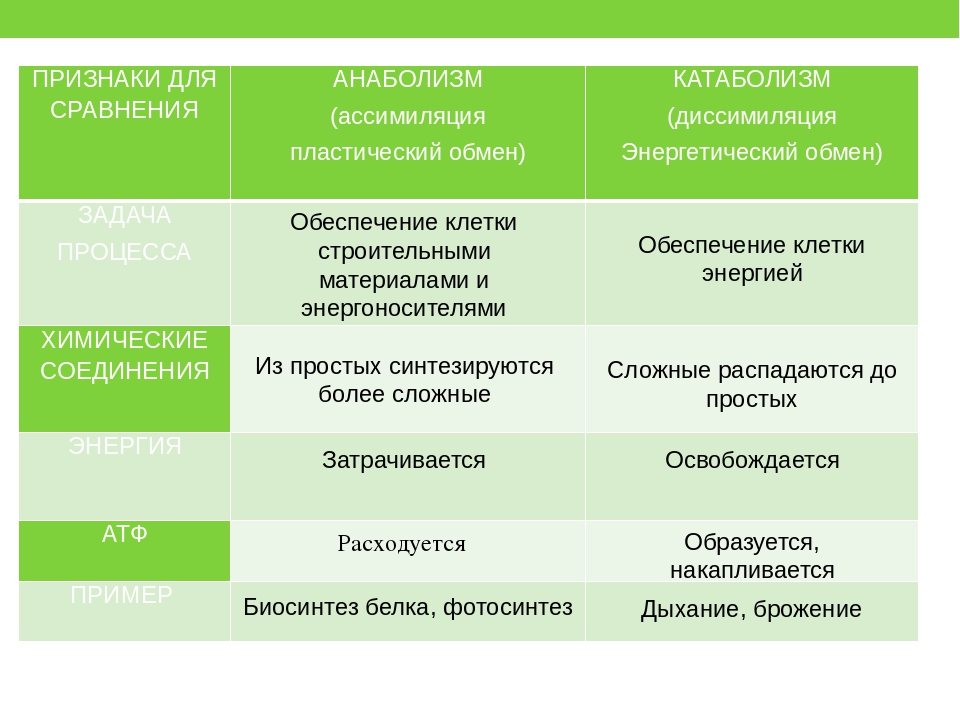 Это, в зависимости от вашего положения, либо банальное, либо неизвестное. Выдвигая на передний план инструментализацию, цель этого эссе — сделать его роль в формировании нашего общего понимания мира немного менее непрозрачной.
Это, в зависимости от вашего положения, либо банальное, либо неизвестное. Выдвигая на передний план инструментализацию, цель этого эссе — сделать его роль в формировании нашего общего понимания мира немного менее непрозрачной.
Метаболические знания производятся флуксометрически — они заключаются в описании и измерении потоков материалов и энергии. Во все более распространенных процессах экологической оценки и рейтинга, в которых человеческая деятельность характеризуется с точки зрения их планетарных «внешних эффектов» углерода или воплощенной энергии, метаболизм является основной истиной анализа.Из всех множества органических метафор, которыми город был обременен историей, метаболизм может в конце концов оказаться наиболее значимым.
ГОРОДСКОЙ МЕТАБОЛИЗМ — ДВА ИЗОБРАЖЕНИЯ
Накагинская капсульная башня. Фото Люка Джонса
Два изображения «городского метаболизма» появились в 1960-х годах, вызванные технологическим кризисом и нанесением ущерба окружающей среде. Для группы архитекторов в Японии потенциал метаболических прочтений города был радикальной дестабилизацией формальной структуры. Метаболизм: предложения для нового урбанизма , представленный группой молодых архитекторов на Всемирной конференции дизайна в Токио в 1960 году, рассматривает метаболизм как движущую силу города, характеризующегося неопределенностью, органическим ростом и нестабильностью. Манифест, содержащий предложения и эссе Киёнори Кикутакэ, Кишо Курокавы и Нобору Кавадзо, представляет собой радикальное преобразование города; преобразован из статической сборки памятников и зданий в гибкий и адаптируемый ландшафт модулей и циркуляционных инфраструктур.Метаболическое пространство мобильно, изменчиво и эмоционально интенсивно, но также связано с природными ландшафтами и циклами.
Для группы архитекторов в Японии потенциал метаболических прочтений города был радикальной дестабилизацией формальной структуры. Метаболизм: предложения для нового урбанизма , представленный группой молодых архитекторов на Всемирной конференции дизайна в Токио в 1960 году, рассматривает метаболизм как движущую силу города, характеризующегося неопределенностью, органическим ростом и нестабильностью. Манифест, содержащий предложения и эссе Киёнори Кикутакэ, Кишо Курокавы и Нобору Кавадзо, представляет собой радикальное преобразование города; преобразован из статической сборки памятников и зданий в гибкий и адаптируемый ландшафт модулей и циркуляционных инфраструктур.Метаболическое пространство мобильно, изменчиво и эмоционально интенсивно, но также связано с природными ландшафтами и циклами.
Городские предложения метаболизма не столько связаны с идентификацией таких потоков самих по себе, сколько с расширением логики потока, неопределенности, квазиорганического роста и адаптации в структуру города. Часто, подобно современной тенденции «подключаемых модулей» в Европе, метаболисты применяют стратегию крупномасштабных мегаструктурных арматур, на которых могут быть размещены более короткоживущие и легко заменяемые компоненты в масштабах комнат и помещений.В Капсульной башне Накагин Курокавы в Симбаси, Токио, возведенной за 30 дней в 1972 году, отдельные мини-квартиры соединены парой бетонных несущих элементов, которые поддерживают их и обеспечивают циркуляцию.
Часто, подобно современной тенденции «подключаемых модулей» в Европе, метаболисты применяют стратегию крупномасштабных мегаструктурных арматур, на которых могут быть размещены более короткоживущие и легко заменяемые компоненты в масштабах комнат и помещений.В Капсульной башне Накагин Курокавы в Симбаси, Токио, возведенной за 30 дней в 1972 году, отдельные мини-квартиры соединены парой бетонных несущих элементов, которые поддерживают их и обеспечивают циркуляцию.
Немногие из реализованных проектов Метаболистов когда-либо реализовали идею непрерывно реконфигурируемой структуры на практике, но по крайней мере такие проекты, как Накагин (или обширная современная схема Кэндзо Танге для Токийского залива), достигают убедительного растворения монументального образа архитектура в состоянии явного движения.
Метаболический эксперимент Санторио, описанный в De Medica Statica, 1615
Стремление группы — изложенное во введении к манифесту — не столько раскрывать метаболизм как основную логику или реальность, сколько воплощать его в реальность на уровне опыта. Для таких архитекторов, как Курокава, динамизм и реконфигурация предлагают городу возврат к своего рода естественному порядку. Здания будут расти, развиваться и реагировать на окружающую среду, как растения на климат.И хотя скорость и адаптируемость современных технологий важны, общество само по себе не является метаболическим — скорее, метаболизм — это случайный способ максимизировать и реализовать присущие ему энергию и динамизм.
Для таких архитекторов, как Курокава, динамизм и реконфигурация предлагают городу возврат к своего рода естественному порядку. Здания будут расти, развиваться и реагировать на окружающую среду, как растения на климат.И хотя скорость и адаптируемость современных технологий важны, общество само по себе не является метаболическим — скорее, метаболизм — это случайный способ максимизировать и реализовать присущие ему энергию и динамизм.
Мы должны, писал Нобору Кавазоэ, «попытаться стимулировать активное метаболическое развитие нашего общества». Или, как утверждал критик и энтузиаст плагинов Рейнер Банхэм в том же духе в выпуске Design Quarterly за 1965 год, «город плагинов должен выглядеть как город плагинов — [люди] должны уметь распознавать его части. и функции, чтобы они могли понять, что он делает с ними, а они с ним.
Представленный пятью годами позже инженером по санитарии Абелем Вольманом в выпуске Scientific American , «Метаболизм города», напротив, представляет собой новую форму анализа продолжающегося кризиса.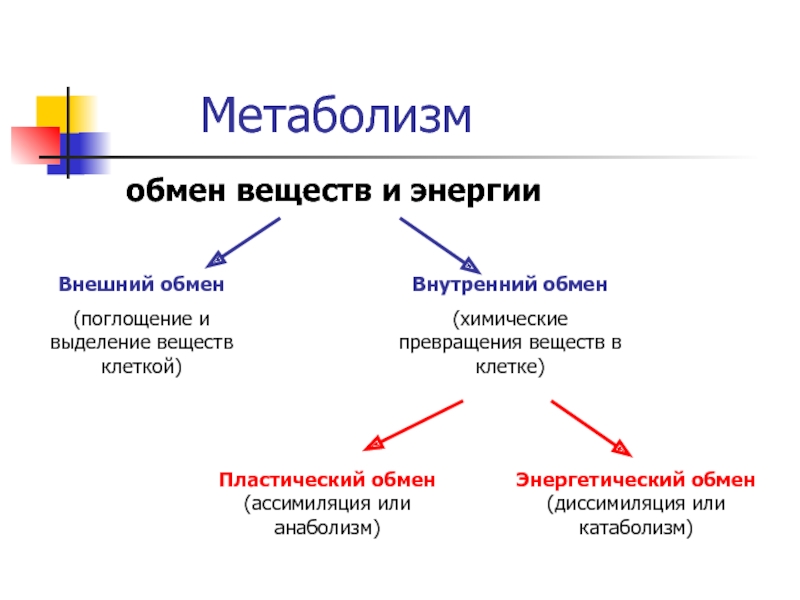
«Метаболические потребности города, — утверждает он, — можно определить как все материалы и товары, необходимые для поддержания жизни жителей города…»
«… Поскольку человек начал понимать, что Земля — это закрытая экологическая система, случайные методы, которые когда-то казались удовлетворительными для удаления отходов, больше не кажутся приемлемыми.У него есть ежедневные свидетельства своих глаз и носа, которые говорят ему, что его планета не может без ограничений ассимилировать необработанные отходы его цивилизации ».
«Минимальный метаболизм»
В наборе графиков и диаграмм Вулман количественно определяет основные входы и выходы, необходимые для гипотетического американского города с миллионным населением, включая воду, продукты питания, первичные энергоносители, такие как нефть, и выходы, например сточные воды, твердые отходы и вредные выбросы. В некотором смысле «Метаболизм города» является естественным продолжением моделей непрерывного гидравлического потока в исходной области Вулмана на более широкую территорию материальных и энергетических потоков.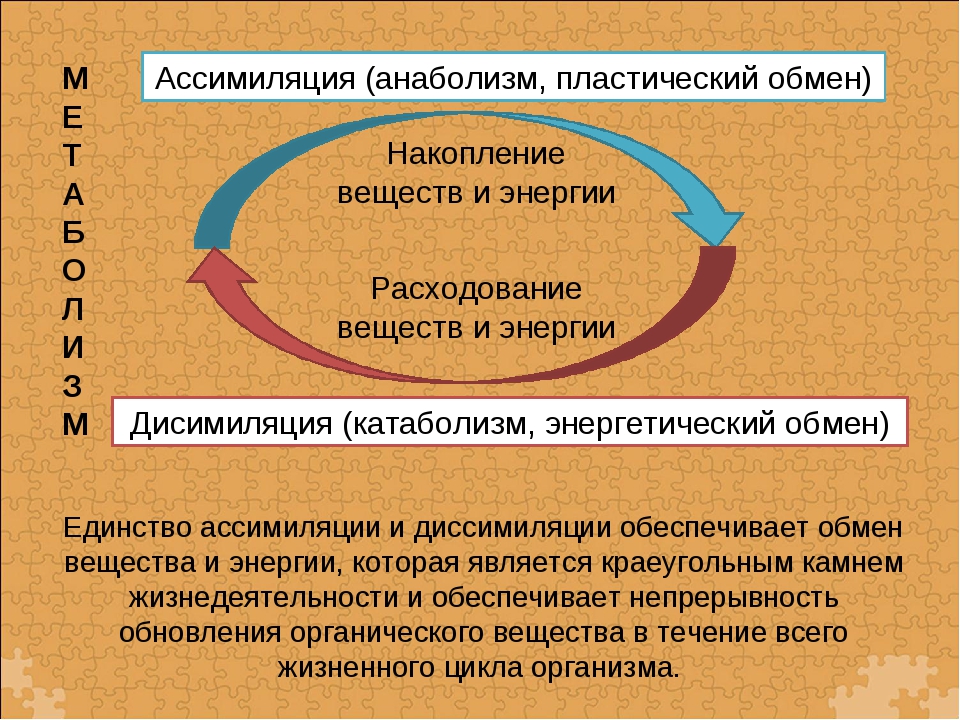 Главный интерес Вулмана — проблема отходов, и его вмешательство является ответом на растущую обеспокоенность общества по поводу загрязнения сточными водами Великих озер и накопления смога над такими городами, как Лос-Анджелес.
Главный интерес Вулмана — проблема отходов, и его вмешательство является ответом на растущую обеспокоенность общества по поводу загрязнения сточными водами Великих озер и накопления смога над такими городами, как Лос-Анджелес.
Между двумя метаболизмами на карту поставлен вопрос масштаба, а также доступности или четкости метаболического процесса. Долговечность архитектуры метаболизма как образа частично объясняется ее убедительной артикуляцией неопределенности или потока на аффективном и узнаваемом уровне.Проект Вулмана, напротив, представляет собой визуализацию в конечном итоге отдаленной целостности. Никто, живущий внутри объекта городского исследования метаболизма, не видит метаболических потоков, в которых они участвуют. Напротив, невидимость этих потоков в повседневной жизни является тем, что мотивирует исследование. Городской метаболизм — это данность. Задача анализа — выявить его, и эта видимость является предпосылкой для регулирования его эффектов.
ТАКТИЧЕСКАЯ НЕОПРЕДЕЛЕННОСТЬ
Методика измерения метаболизма в процессе работы, от Фрэнсиса Бенедикта Человеческая жизнеспособность и эффективность , 1919
История городской теории полна органических метафор. Во Франции восемнадцатого века сформировалось представление о городе как о физиологическом, которое сохраняется в банальных словах сегодняшнего дня — описании главных улиц как артерий или городской текстуры как ткани. Если в городе есть тело, то оно может быть в грубом или хрупком здоровье. Лечение, как при османизации Парижа, может быть хирургическим. В более поздних моделях город стал не столько телом, сколько органической клеткой или единицей — ограниченным, правильного размера, внутренне интегрированным и саморегулирующимся, непреодолимым перед лицом грубых механизмов вмешательства; или продукт квазиестественных эволюционных сил, формирующихся в ответ на изменение ландшафта и внешних требований.В послевоенной кибернетике городской организм — это своего рода неврология, самоуправляемая и самосигнальная, выстраивающая петли обратной связи и сетевые отношения все большей абстракции и масштаба. Аналогии могут быть взаимно дополняющими или несовместимыми — риторика организма, безусловно, использовалась как яростная поддержка модернистского проекта, а также его осуждение.
Во Франции восемнадцатого века сформировалось представление о городе как о физиологическом, которое сохраняется в банальных словах сегодняшнего дня — описании главных улиц как артерий или городской текстуры как ткани. Если в городе есть тело, то оно может быть в грубом или хрупком здоровье. Лечение, как при османизации Парижа, может быть хирургическим. В более поздних моделях город стал не столько телом, сколько органической клеткой или единицей — ограниченным, правильного размера, внутренне интегрированным и саморегулирующимся, непреодолимым перед лицом грубых механизмов вмешательства; или продукт квазиестественных эволюционных сил, формирующихся в ответ на изменение ландшафта и внешних требований.В послевоенной кибернетике городской организм — это своего рода неврология, самоуправляемая и самосигнальная, выстраивающая петли обратной связи и сетевые отношения все большей абстракции и масштаба. Аналогии могут быть взаимно дополняющими или несовместимыми — риторика организма, безусловно, использовалась как яростная поддержка модернистского проекта, а также его осуждение.
Городские организмы предоставляют когнитивные карты для вмешательства. Если город — это тело, то знание его анатомии будет происходить через рассечение.Если это больше похоже на экосистему, то методы экологической съемки могут быть более подходящими. В неврологической системе построение графиков и моделирование сетей более высокого уровня, по-видимому, наиболее вероятно улавливает основную динамику в действии.
Аналоги тел или клеток, по крайней мере, частично структурные; напротив, метаболизм в основном бесформенный. В том смысле, что все другие организмы что-то говорят о том, что такое объект, метаболизм определяет только то, что он делает. Там, где другие органические аналогии идентифицируют системы гомеостатической саморегуляции (или даже интеллекта), метаболизм, по сути, тупой.Городской метаболизм лишен всяких претензий — в смысле культуры, ценности, значения, значения или даже намерения. Это пищеварение, а не намерение. У него нет цели, нет инстинкта, нет цели. Идентификация человеческих процессов как метаболических не имеет никакого отношения к балансу или устойчивости; напротив, он указывает на отсутствие этих форм регулирования.
Расширение лимфатических сосудов у плода человека, от Франца Крайбеля, Руководство по эмбриологии человека , 1910
Метаболизм как метод анализа является сильно, даже грубо, восстановительным.Некоторые из самых ранних метаболических экспериментов проводил венецианский врач Санторио Санторио. Выявив массовый дисбаланс между едой и напитками, которые он потреблял, и количеством выделяемых им фекалий и мочи, он решил измерить потерю массы тела за счет потоотделения, сидя на огромных безбилетных весах в течение нескольких часов после еды. отслеживать его постепенное изменение массы.
Эксперимент дает минимальное определение метаболизма как измерения потоков в системе и из нее, в данном случае частично за счет принципа баланса массы.Эти потоки конкретно познаваемы и поддаются количественной оценке — где внутренняя часть системы (или тела) может быть только предположена. Граница метаболизма — это место, где собирается информация — ее внутренняя часть, эпистемически или тактически, не определена.
Эта последняя часть важна — проект метаболического анализа повседневной жизни можно рассматривать как основанный на определенном уровне идеи создания петли обратной связи, которой раньше не было; между деятельностью человека, от покупки продуктов до проектирования зданий, и их широкомасштабным воздействием на планету.Такие показатели, как воплощенный углерод или экологический след, читаются как попытки произвести системную осведомленность внутри систем, которые не являются разумными по своей сути.
СРЕДИ ПОТОКОВ
То, что поставлено на карту для видимости или прозрачности метаболического потока или действий в масштабе человеческой жизни и взаимодействия, стало ясно только в последние десятилетия, когда центр экологического менеджмента и реформ сместился с центров производство в области потребления.
Во времена Уолмана основным направлением экологического проектирования и управления было то, что было названо «концом трубы» — основная работа и влияние крупномасштабных инфраструктур и процессов.В последние несколько десятилетий произошел вековой сдвиг, в котором сфера потребления — «сторона спроса» — становится все более целевой.
Для того, чтобы этот сдвиг стал возможным, необходимо установить связь между флуксометрией рассматриваемых метаболических процессов — городами, фабриками, отраслями, регионами — и масштабами человеческого взаимодействия. Продукты, здания, проекты должны быть представлены как fluxomes — то есть как наборы или связки потоков; каждый как крошечный толчок, так или иначе, в сдвигах и нарастаниях потока планетарного масштаба.
ОТ ЖИЗНИ К ПЛАТФОРМАМ
Этапы оценки жизненного цикла (ISO 14040)
В современной практике экологической оценки определение деятельности человека в терминах метаболизма является хорошо понятным процессом. Потоки материалов, энергии, тепла, отходов, пара на заводе и за его пределы или процесс транспортировки транспортных средств в виде потока углеводородов, превращающегося в выбросы, могут быть напрямую количественно определены и охарактеризованы.
Но метаболическая характеристика объекта — обуви или здания — является более сложной проблемой.Объекты перемещаются, и они не остаются самими собой, а собираются, разбираются, расходуются и так далее. Они движутся в пространстве и времени и являются участниками последовательности различных процессов, когда их очищают, собирают, продают, используют и выбрасывают.
Начиная с 1980-х и 1990-х годов как механизм оценки воздействия проектов и инвестиций, оценка жизненного цикла стала доминирующей процедурой для формирования экологических объектов, то есть объектов, сформированных вокруг визуализации их внешних воздействий на окружающую среду.Принцип этих упражнений, как хорошо известно, состоит в том, чтобы дать структурную разбивку внешних эффектов по стадиям. Эти стадии сами по себе являются фракциями или абстракциями метаболических анализов процессов. Потоки, связанные с транспортным средством, подразделяются и привязаны к объектам, которые они доставляют; фабрики используются для характеристики продуктов, которые они создают. Некоторые из этих процессов исследуются напрямую, другие представляют собой общие данные из общих баз данных и программных платформ.
«Границы системы» жизненного цикла расширяются на периоды времени и включают в себя сценарии будущих показателей, например, энергозатраты на потребление при использовании или углеродоемкость рециркуляции или утилизации.Энергетические затраты на переработку элемента должны включать стоимость его жизненного цикла сегодня, даже если этот процесс может потребоваться в будущем энергетическом режиме весьма неопределенного характера. Сколько углерода будет выделять электричество, используемое для переработки продукта, через тридцать лет? Какие механизмы безопасного удаления или повторного использования будут доступны? Эти процессы композиции и разложения, генерации и проецирования лежат в основе очевидных определений воздействия на окружающую среду, таких как углеродоемкость, экологический след и воплощенная энергия.
Цепочки непредвиденных обстоятельств в техносфере
Жизненные циклы все чаще становятся протоколом для более сложных систем оценки. В экологических декларациях продуктов (EPD) они прилагаются к продуктам и товарам, которые, в свою очередь, интегрируются в более масштабные оценки. LCA в масштабе здания обязательно объединяет целый набор LCA в масштабе продукта для его различных элементов. В недавних проектах была предпринята попытка интегрировать их непосредственно в программное обеспечение САПР и информационного моделирования зданий (BIM) — например, проект «углеродный калькулятор» EC3, осуществляемый Skanska, Microsoft и другими.Платформы встроены на обоих концах процесса — при определении промышленных процессов как общих данных (тысячи из которых содержатся в широко используемой базе данных Ecoinvent) или при использовании EPD (или все чаще их машиночитаемой формы — ILCD) сами по сравнительным базам данных.
Теоретически цель этих процессов в совокупности состояла бы в том, чтобы позволить городу или экономике ясно увидеть собственное воздействие на окружающую среду; для данного элемента — кирпича, кондиционера — соответствующее увеличение содержания углерода в атмосфере.Объекты повседневной жизни постепенно превращаются в их симулякры.
ЛАНДШАФТ ПОТОКОВ
Запутанные потоки в производстве стали и бетона
Мир повсеместного жизненного цикла построен странным циклом, в котором объективный метаболический сигнал и культурная система, которую он призван исправлять, постоянно перезаписывают один еще один.
Процессы в техносфере часто переплетаются или складываются вместе, и их можно разделить только путем интерпретации.На птицеферме могут выращиваться цыплята, яйца, куриный помет. Доменная печь может производить чугун, доменный шлак и тепло. Не существует неизбежного способа определить, кто из них отвечает в системе за внешние эффекты процесса, из которого они происходят. На каком основании, например, выбросы углекислого газа в процессе потребления железа, угля и электричества — и получения шлака, чугуна и тепла — распределяются между тем или иным производством? Какой из «продуктов» их вызывает?
В строительстве обычным способом снижения выбросов углерода из бетона является замена некоторой части связующего портландцемента измельченным гранулированным шлаком при производстве стали.Цемент является наиболее углеродоемким ингредиентом бетона и производит около 0,9 тонны CO2 на тонну цемента, большая часть из которых возникает в процессе прокаливания. Шлак получают при выплавке стали из руды из расчета около 150 кг на тонну стали, в процессе, который производит от 1,9 до 2,3 тонн CO2 на тонну чугуна. С чисто метаболической точки зрения шлак, чугун, диоксид углерода и тепло являются выходами процесса, но диоксид углерода и другие внешние факторы связаны с жизненным циклом стали, позволяя шлаку улетучиваться в качестве чистого побочного продукта.Если бы в каком-то своеобразном зеркальном мире целью процесса плавки было производство шлака, а не стали, он выбрасывал бы 12-15 тонн CO2 на тонну GGBS.
Современные передовые практики дают несколько ответов на эти вопросы. Вы можете распределить внешние эффекты по экономической стоимости выпуска или их относительной массе. Вы можете отделять отходы от продуктов по категориям — исходя из того, что в процессе производится специально, а не как «побочный продукт». Вы можете решить, что бы люди делали вместо этого, если бы продукта не существовало; о том, что они прекратили бы делать, если бы оно стало дешевле или более доступным.Но в любом из этих случаев очевидный внешний объективный сигнал и объект анализа должны быть перекалиброваны в рамках тех же культурных систем и предположений, которые они предлагают просветить, а в некоторых случаях круговыми способами. Связь между метаболизмом как реальностью и нашей частью опосредована способами, которые в конечном итоге затемняют сигнал.
СИСТЕМЫ УПРАВЛЕНИЯ
По мнению эколога Питера Хаффа, эта проблема является неотъемлемой частью человеческой «техносферы». Метаболические потоки, действия и выбор человека — это одна и та же система, но с совершенно несовместимыми внешними проявлениями и переменными.Та же грубость зерна, которая делает лист в одном масштабе клеточным ландшафтом, а в другом — дискретным объектом, указывает на разделение «слоев» в сложных системах. То, что можно визуализировать в изображении региональных или континентальных величин как одного набора переменных, будет отображаться в масштабе повседневной человеческой деятельности как нечто совершенно иное — и хотя планетарная катастрофа проявляется в самом верхнем слое, механизмы контроля, такие как они есть, все на одном ниже.Люди, как заметил Хафф, «являются компонентами более крупной сферы, которую они не проектировали, не понимают, не контролируют и из которой они не могут убежать».
Потоки в одном процессе
В масштабе городов, стран и более крупных процессов мы видим потоки, но в масштабе человеческой деятельности эти потоки естественным образом скрыты. Мы визуализируем и взаимодействуем не с потоками, а с объектами — зданиями, товарами, продуктами и т. Д. — внутри которых эти планетные движения больше не видимы в естественном смысле.Не только невидимы переменные верхнего слоя, но и неясна связь объекта с ними.
Между нашим собственным слоем и слоями ниже, как заметил Хафф, мы обязательно используем микроскопы, центрифуги, зонды и манипуляторы различных типов, чтобы вмешиваться в набор переменных, которые иначе были бы недоступны.
ФЛЮКСОМЕТРИЧЕСКОЕ УПРАВЛЕНИЕ
Если запутанные кризисы чрезмерной добычи, загрязнения и выбросов углерода являются результатом неупорядоченной материальной культуры, то их решением может быть некая форма управления.Найти какой-то способ соединения слоев — сформировать своего рода техносоциальный микроскоп — является предпосылкой для целостного регулирования и самокоррекции материальной системы. Управление стало возможным благодаря технологиям. Современное государство сформировалось на основе таких инструментов — от землеустройства до учета, переписи населения и видеонаблюдения.
Если целью метаболизма является восстановление отношений между городом и его геобиофизическими внутренними районами, это обязательно будет происходить по определенной схеме.Самоосознание постантропоценового города будет поддерживаться флуксометрикой и воспроизводить конкретные структуры и качества этой техники. Метаболизм соединяет мир вверху с миром внизу посредством двойного перевода. Во-первых, знакомые объекты попадают в безличный ландшафт химических потоков. Затем эти потоки подразделяются и перераспределяются между вызвавшими их человеческими процессами — как «воздействия на окружающую среду». Несовместимость этих систем знания — поля потоков и дискретного каталога объектов — в какой-то степени неизбежно приводит к случайным сбоям в системе экологической ценности.Гибкость метаболизма как концепция — это то, что связывает внутреннюю часть человеческой культуры с планетарной внешностью, а затем обратно.
Натуралистическая аналогия в применении к человеческим процессам, таким образом, порождает неоднозначное отношение к самой природе. Как определение процессов через химический поток, универсально применяемое к антропогенной и нечеловеческой деятельности, идея метаболизма в некотором смысле отменяет качественное различие между одним и другим. То, что остается, скорее, между человеком и нечеловеческим, биосферой и техносферой, — это необходимое, но чисто догматическое разделение между двумя эквивалентными полями.Это разделение на самом фундаментальном уровне является «реальностью» самого метаболизма, что позволяет ему собирать информацию.
По мере того как экологические индексы становятся все более встроенной корректирующей моделью человеческого дизайна и производства, становится важным определить формы и методы, с помощью которых такие системы создаются и обслуживаются. В мире повсеместной экологической индексации или расчета углерода наша общая реальность становится флюксометрикой человеческих процессов.Чем больше мы учитываем при проектировании наших городов расчет воздействия на окружающую среду как социального или технического блага, тем больше мы становимся гражданами fluxopia . При визуализации нашего общего метаболизма в решающих масштабах и местах объективность исходного сигнала постепенно теряется. Мы можем осознать свое собственное воздействие, только сделав его полностью неузнаваемым; мир изменился, так что он в конечном итоге может оставаться более или менее самим собой. Именно эту медленную, рекурсивную самокоррекцию мы можем ожидать снова и снова читать в развивающихся структурах планетарного самосознания.
Люк Джонс
Люк Джонс является партнером Heat Island в Лондоне и был научным сотрудником по дистанционным исследованиям программы терраформирования в цикле 2020 года. Он ведет подкаст «О зданиях и городах».
Гемоглобин и миоглобин — Медицинская биохимия Страница
Миоглобин
Миоглобин и гемоглобин — это гемопротеины, физиологическое значение которых в основном связано с их способностью связывать молекулярный кислород.Миоглобин — это мономерный гем-белок, обнаруживаемый в основном в мышечной ткани, где он служит местом внутриклеточного хранения кислорода. В периоды кислородного голодания оксимиоглобин выделяет связанный кислород, который затем используется для метаболических целей.
Третичная структура миоглобина — это структура типичного водорастворимого глобулярного белка. Его вторичная структура необычна тем, что содержит очень высокую долю (75%) α-спиральной вторичной структуры. Полипептид миоглобина состоит из 8 отдельных правосторонних α-спиралей, обозначенных от А до Н, которые соединены короткими неспиральными участками.Аминокислотные R-группы, упакованные внутрь молекулы, имеют преимущественно гидрофобный характер, в то время как те, которые выставлены на поверхности молекулы, обычно гидрофильны, что делает молекулу относительно водорастворимой.
Структура миоглоина с гемомКаждая молекула миоглобина содержит одну простетическую группу гема, вставленную в гидрофобную щель в белке. Каждый остаток гема содержит один центрально связанный атом железа с координационной связью, который обычно находится в степени окисления Fe 2+ (двухвалентное железо).Кислород, переносимый гемепротеинами, связан непосредственно с атомом двухвалентного железа простетической группы гема. Гидрофобные взаимодействия между тетрапиррольным кольцом и R-группами гидрофобной аминокислоты внутри расщелины в белке сильно стабилизируют конъюгат гем-белка. Кроме того, атом азота из группы R гистидина, расположенный над плоскостью гемового кольца, координирован с атомом железа, дополнительно стабилизируя взаимодействие между гемом и белком. В оксимиоглобине оставшийся участок связывания на атоме железа (шестая координата) занят кислородом, связывание которого стабилизируется вторым остатком гистидина.
Гемоглобин
Гемоглобин взрослых представляет собой [α (2): β (2)] тетрамерный гемепротеин, обнаруженный в эритроцитах, где он отвечает за связывание кислорода в легких и транспортировку связанного кислорода по всему телу, где он используется в аэробных метаболических путях.
Структура гемоглобинаОписание различных типов тетрамеров гемоглобина см. Ниже в разделе «Гены гемоглобина». Каждая субъединица тетрамера гемоглобина имеет простетическую группу гема, идентичную описанной для миоглобина.Обычные пептидные субъединицы обозначаются α, β, γ и δ, которые образуют наиболее часто встречающиеся функциональные гемоглобины.
Хотя вторичная и третичная структуры различных субъединиц гемоглобина схожи, отражая обширную гомологию в аминокислотном составе, различия в аминокислотном составе, которые действительно существуют, приводят к заметным различиям в свойствах гемоглобина переносить кислород. Кроме того, четвертичная структура гемоглобина приводит к физиологически важным аллостерическим взаимодействиям между субъединицами — свойству, отсутствующему у мономерного миоглобина, который в остальном очень похож на α-субъединицу гемоглобина.
Сравнение кислородсвязывающих свойств миоглобина и гемоглобина иллюстрирует аллостерические свойства гемоглобина, обусловленные его четвертичной структурой, и дифференцирует кислородсвязывающие свойства гемоглобина от миоглобина. Кривая связывания кислорода с гемоглобином является сигмоидальной, типичной для аллостерических белков, в которых субстрат, в данном случае кислород, является положительным гомотропным эффектором. Когда кислород связывается с первой субъединицей дезоксигемоглобина, он увеличивает сродство остальных субъединиц к кислороду.По мере того как дополнительный кислород связывается со второй и третьей субъединицами, связывание кислорода постепенно усиливается, так что при напряжении кислорода в альвеолах легких гемоглобин полностью насыщается кислородом. По мере того, как оксигемоглобин циркулирует в деоксигенированной ткани, кислород постепенно разгружается, и сродство гемоглобина к кислороду снижается. Таким образом, при самом низком напряжении кислорода, обнаруженном в очень активных тканях, сродство связывания гемоглобина с кислородом очень низкое, что обеспечивает максимальную доставку кислорода к ткани.Напротив, кривая связывания кислорода для миоглобина имеет гиперболический характер, что указывает на отсутствие аллостерических взаимодействий в этом процессе.
Кривые насыщения кислородом миоглобина и гемоглобина. Кривая насыщения миоглобина показывает типичное быстрое зависимое от концентрации кислорода насыщение этого мономерного связывающего кислород белка. Две другие кривые показывают типичные сигмоидальные кривые насыщения для кооперативного связывания кислорода, проявляемого фетальным гемоглобином (HbF) и взрослым гемоглобином (HbA).На диаграмме также указаны типичные концентрации кислорода в периферических тканях и легких. Обратите внимание, что в то время как миоглобин может быть полностью насыщен кислородом в тканях, гемоглобину требуется гораздо более высокое напряжение кислорода, чтобы стать полностью насыщенным, что происходит только в легких. Положение насыщения HbF слева от HbA (то есть при более низком давлении кислорода) отражает тот факт, что гемоглобин плода связывает кислород с более высоким сродством, чем гемоглобин взрослого, и это сделано для того, чтобы плод мог получать кислород из материнского кровообращения.Аллостерические кислородсвязывающие свойства гемоглобина возникают непосредственно в результате взаимодействия кислорода с атомом железа простетических групп гема и результирующего воздействия этих взаимодействий на четвертичную структуру белка. Когда кислород связывается с атомом железа дезоксигемоглобина, он втягивает атом железа в плоскость гема. Поскольку железо также связано с гистидином F8, этот остаток также притягивается к плоскости гемового кольца. Конформационное изменение гистидина F8 передается по всей пептидной цепи, что приводит к значительному изменению третичной структуры всей субъединицы.Конформационные изменения на поверхности субъединицы приводят к новому набору связывающих взаимодействий между соседними субъединицами. Последние изменения включают разрушение солевых мостиков и образование новых водородных связей и новых гидрофобных взаимодействий, все из которых вносят вклад в новую четвертичную структуру.
Последние изменения во взаимодействии субъединиц передаются с поверхности в гем-связывающий карман второй дезокси-субъединицы и приводят к более легкому доступу кислорода к атому железа второго гема и, таким образом, большему сродству молекулы гемоглобина к вторая молекула кислорода.Третичная конфигурация низкого сродства, деоксигенированного гемоглобина (Hb) известна как напряженное (T) состояние. И наоборот, четвертичная структура полностью оксигенированной формы гемоглобина с высоким сродством (HbO 2 ) известна как расслабленное (R) состояние.
В контексте сродства гемоглобина к кислороду существует четыре основных регулятора, каждый из которых оказывает негативное влияние. Это CO 2 , ион водорода (H + ), хлорид-ион (Cl — ) и 2,3-бисфосфоглицерат (2,3-БПГ, а также просто БПГ).В некоторых более старых текстах 2,3-BPG сокращено до 2,3-DPB. Хотя они могут влиять на связывание O 2 независимо друг от друга, CO 2 , H + и Cl — в основном действуют как следствие друг друга на сродство гемоглобина к O 2 . Сначала мы рассмотрим перенос O 2 из легких в ткани.
В среде легких с высоким содержанием O 2 (высокий pO 2 ) достаточно O 2 для преодоления ингибирующей природы Т-состояния.Во время индуцированного связыванием O 2 изменения от Т-формы к R-форме несколько боковых аминокислотных групп на поверхности субъединиц гемоглобина будут диссоциировать протоны, как показано в уравнении ниже. Эта диссоциация протонов играет важную роль в истечении CO 2 , который поступает из тканей (см. Ниже). Однако из-за высокого pO 2 pH крови в легких (≈7,4–7,5) недостаточно низок, чтобы оказывать негативное влияние на связывание гемоглобина O 2 .Когда оксигемоглобин достигает тканей, pO 2 достаточно низкое, а также pH (≈7,2), что благоприятствует T-состоянию и высвобождению O 2 .
4 O 2 + Hb ↔ nH + + Hb (O 2 ) 4
Если мы теперь рассмотрим, что происходит в тканях, можно увидеть, как CO 2 , H + и Cl — оказывают отрицательное влияние на связывание гемоглобина O 2 .Метаболические клетки производят CO 2 , который диффундирует в кровь и попадает в циркулирующие эритроциты (RBC). В эритроцитах CO 2 быстро превращается в угольную кислоту под действием карбоангидразы, как показано в уравнении ниже:
CO 2 + H 2 O ↔ H 2 CO 3 ↔ H + + HCO 3 —
Ион бикарбоната, образующийся в этой реакции диссоциации, диффундирует из эритроцитов и переносится с кровью в легкие.Этот эффективный процесс переноса CO 2 называется изогидным переносом. Таким образом, примерно 80% CO 2 , продуцируемого метаболизирующими клетками, транспортируется в легкие. Небольшой процент CO 2 переносится с кровью в виде растворенного газа. В тканях H + , диссоциированный от угольной кислоты, забуферен гемоглобином, который оказывает негативное влияние на связывание O 2 , вызывая высвобождение в тканях. Как указано выше, в легких высокий pO 2 обеспечивает эффективное связывание O 2 гемоглобином, что приводит к переходу из состояния T в состояние R и высвобождению протонов.Протоны соединяются с бикарбонатом, поступающим из тканей, образуя угольную кислоту, которая затем попадает в эритроциты. За счет обращения карбоангидразной реакции образуются CO 2 и H 2 O. CO 2 диффундирует из крови в альвеолы легких и высвобождается по истечении срока годности.
Помимо изогидрического транспорта, до 15% CO 2 транспортируется в легкие, связавшись с N-концевыми аминогруппами Т-формы гемоглобина.Эта реакция, изображенная ниже, образует так называемый карбаминогемоглобин . Как указано, эта реакция также производит H + , тем самым снижая pH в тканях, где концентрация CO 2 высока. Образование H + приводит к высвобождению связанного O 2 в окружающие ткани. В легких высокое содержание O 2 приводит к связыванию O 2 с гемоглобином с одновременным высвобождением H + . Высвободившиеся протоны затем способствуют диссоциации карбамино с образованием CO 2 , который затем высвобождается с истечением срока годности.
CO 2 + Hb-NH 2 ↔ H + + Hb-NH-COO —
Как показывает приведенное выше обсуждение, конформация гемоглобина и его связывание с кислородом чувствительны к концентрации ионов водорода. Эти эффекты концентрации ионов водорода ответственны за хорошо известный эффект Бора , в котором увеличение концентрации ионов водорода уменьшает количество кислорода, связанного гемоглобином при любой концентрации кислорода (парциальном давлении).Наряду с диффузией бикарбоната из эритроцитов в ткани должно происходить движение ионов в эритроциты для поддержания электрической нейтральности. Это роль Cl —, которая называется сдвигом хлорида . Таким образом, Cl — играет важную роль в производстве и диффузии бикарбоната и, таким образом, также отрицательно влияет на связывание O 2 с гемоглобином.
Представление переноса CO 2 из тканей в кровь с доставкой O 2 в ткани. CO 2 , образующийся в результате метаболических процессов в тканях, диффундирует в кровь. Около 2–5% CO 2 остается в крови в виде растворенного газа. Большая часть CO 2 улавливается эритроцитами, где он образует комплекс с водой под действием карбоангидразы (CA), образующей угольную кислоту. Угольная кислота ионизируется, и высвободившийся ион бикарбоната (HCO 3 –) транспортируется в кровь через транспортер SCL4A1 (также известный как анионообменник 1, AE1) в обмен на ион Cl–.Этот процесс называется хлоридным сдвигом. Бикарбонат в крови составляет от 45% до 50% транспортной формы CO 2 . Около 5% –15% ткани CO 2 транспортируется ковалентно связанным с гемоглобином в виде карбаминогемоглобина. Противоположный процесс происходит, когда O 2 поступает из альвеол легких, а CO 2 удаляется.Метгемоглобин
Окисление двухвалентного (Fe 2+ ) железа до степени окисления трехвалентного (Fe 3+ ) делает глобиновый мономер неспособным к нормальному связыванию кислорода.Когда одна или несколько молекул гемового железа находятся в трехвалентном состоянии, гемоглобин называется , метгемоглобин . Образование активных форм кислорода (АФК), супероксид-аниона, может происходить в эритроцитах (красных кровяных тельцах, эритроцитах) в результате окисления двухвалентного железа (Fe 2+ ) до трехвалентного железа (Fe 3) O 2 + ) в гемоглобине. Следствием последней реакции является образование метгемоглобина. Поскольку гемоглобин является гетеротетрамером, и каждая субъединица содержит железо Fe 2+ в своем геме, существует вероятность того, что в метгемоглобине могут присутствовать несколько железа Fe 3+ .Форма железа Fe 3+ не связывает O 2 , однако присутствие по крайней мере одного Fe 3+ в тетрамере гемоглобина приводит к усиленному связыванию O 2 с оставшимся Fe 2 + утюгов, вызывающих снижение доставки O 2 к тканям с потенциалом цианоза. Нормальный дневной уровень метгемоглобина колеблется в пределах 0,5–3%.
Трехвалентное железо в метгемоглобине восстанавливается до двухвалентного под действием НАДН-требующего фермента, метгемоглобинредуктазы (цитохром b 5 редуктаза 3: CYB5R3).Белок, кодируемый CYB5R3, растворим в эритроцитах, но связан с мембраной в клетках других типов. Мембраносвязанная форма связана с мембранами эндоплазматического ретикулума (ER), где она участвует в удлинении и десатурации жирных кислот, в синтезе холестерина и в метаболизме ксенобиотиков. Люди экспрессируют четыре гена цитохрома b 5 редуктазы, CYB5R1, CYB5R2, CYB5R3 и CYB5R4.
НАДФН, продуцируемый пентозофосфатным путем (PPP), и антиоксидантный глутатион (GSH) необходимы для постоянного удаления ROS из эритроцита.
Окись углерода, CO
Окись углерода (CO) также координированно связывается с атомами железа гема в двухвалентном состоянии (Fe 2+ ) в гемоглобине аналогично кислороду. Однако связывание CO с гемовым железом намного сильнее (> 200 раз), чем у O 2 . Когда CO связывается с гемоглобином, образующийся белок называется карбоксигемоглобином (HbCO). Помимо более высокого сродства CO по сравнению с O 2 к двухвалентному железу в гемоглобине, результат связывания CO по крайней мере с одним мономером глобина вызывает более высокое сродство остальных субъединиц к O 2 .Этот эффект способствует снижению высвобождения O 2 в тканях. Эти комбинированные эффекты предпочтительного связывания CO с гемовым железом в значительной степени ответственны за удушье, которое возникает в результате отравления монооксидом углерода.
Роль 2,3-бисфосфоглицерата (2,3-BPG)
Соединение 2,3-бисфосфоглицерат (2,3-BPG), полученное из промежуточного гликолитического соединения 1,3-бисфосфоглицерата, является мощным аллостерическим эффектором на свойства связывания кислорода гемоглобина.Путь синтеза 2,3BPG эритроцитов называется шунтом Рапопорта – Люберинга и схематически показан на рисунке ниже.
Бифункциональный фермент, бисфосфоглицератмутаза (кодируемый геном BPGM) переводит часть 1,3-бисфосфоглицерата из гликолитического пути в 2,3-бисфосфоглицерат за счет действия синтазной функции фермента. Превращение 2,3-BPG в 3-фосфоглицерат катализируется функцией фосфатазы BPGM.
Путь синтеза 2,3-бисфосфоглицерата (2,3-BPG) в эритроцитах. Синтез 2,3-BPG представляет собой основной путь реакции потребления глюкозы в эритроцитах. Синтез 2,3-BPG в эритроцитах имеет решающее значение для контроля сродства гемоглобина к кислороду. Обратите внимание, что когда глюкоза окисляется этим путем, эритроцит теряет способность получать 2 моля АТФ от гликолитического окисления 1,3-BPG до 3-фосфоглицерата через реакцию фосфоглицераткиназы.В деоксигенированной конформации Т в центре тетрамера гемоглобина образуется полость, способная связывать 2,3-БПГ.Отдельная молекула 2,3-БПГ может занимать эту полость, тем самым стабилизируя Т-состояние. Взаимодействие 2,3-BPG с дезоксигемоглобином происходит через солевые мостики с остатками Lys и His в субъединицах β-глобина тетрамера гемоглобина. И наоборот, когда 2,3-BPG недоступен или не связан в центральной полости, Hb может легче связывать кислород (образуя HbO 2 ). Таким образом, как и повышенная концентрация ионов водорода (H + ), повышенная концентрация 2,3-BPG способствует преобразованию R-формы Hb в T-форму Hb и снижает количество кислорода, связанного Hb при любой концентрации кислорода.Когда давление кислорода достаточно высокое, например, в альвеолах легких, связывание моля O 2 вызывает переход T-to-R, который вызывает коллапс связующего кармана 2,3-BPG и 2 , 3-BPG удаляется, позволяя остальным мономерам глобинового белка связывать O 2 .
Известно, что молекулы гемоглобина, различающиеся по составу субъединиц, обладают разными свойствами связывания 2,3-BPG с соответственно разными аллостерическими ответами на 2,3-BPG.Например, HbF (эмбриональная форма гемоглобина) состоит из двух α-белков и двух γ-белков. Хотя белки γ-глобина и β-глобина на 72% идентичны на аминокислотном уровне, существует одно существенное аминокислотное различие в связывающем кармане 2,3-BPG. В белке -глобине His 143 заменен на Ser в сайте связывания 2,3-BPG. Эффект этой аминокислотной замены заключается в потере двух положительных зарядов (по одному от каждой γ-субъединицы относительно β-субъединиц). В результате 2,3-BPG гораздо меньше связывается с HbF, чем с HbA (взрослой формой гемоглобина).Следствием этого является то, что HbF у плода связывает кислород с большей аффинностью, чем HbA матери, тем самым обеспечивая плоду преимущественный доступ к кислороду, переносимому кровеносной системой матери.
Гены гемоглобина
Белки α- и β-глобина, содержащиеся в функциональных тетрамерах гемоглобина, происходят из кластеров генов. Гены α-глобина находятся на хромосоме 16, а гены β-глобина находятся на хромосоме 11. Оба кластера генов содержат не только основные гены взрослого человека, α и β, но и другие экспрессируемые последовательности, которые используются на разных стадиях развития.Ориентация генов в обоих кластерах находится в одном направлении от 5 ‘к 3’ с наиболее ранними экспрессируемыми генами на 5 ‘конце обоих кластеров. Помимо функциональных генов, оба кластера содержат нефункциональные псевдогены.
Хромосомная структура кластеров генов α- и β-глобина на хромосомах 16 и 11 соответственно. Ориентация генов с 5 ‘на 3′ на каждой хромосоме также отражает время развития их экспрессии с 5’-самыми ранними генами.Гены ζ (дзета) и ε (эпсилон) являются эмбриональными генами в каждом кластере. Гены с обозначением Ψ (psi) представляют собой псевдогены.Синтез гемоглобина начинается в первые несколько недель эмбрионального развития в желточном мешке. Главный гемоглобин на этой стадии развития представляет собой тетрамер, состоящий из двух дзета (ζ) цепей, кодируемых в α-кластере, и двух эпсилон (ε) цепей из β-кластера. К 6-8 неделям беременности экспрессия этой версии гемоглобина резко снижается, что совпадает с изменением синтеза гемоглобина из желточного мешка в печень.Экспрессия из кластера α состоит из идентичных белков из генов α1 и α2. Экспрессия этих генов в α-кластере сохраняется на протяжении всей жизни.
Внутри кластера β-глобина есть дополнительный набор генов, гены β-глобина плода, идентифицированные как гены гамма (γ). Два гена плода, называемые Gγ и Aγ, происхождение которых связано с единственной аминокислотной разницей между двумя генами плода: глицин в Gγ и аланин в Aγ в положении 136. Эти гены γ плода экспрессируются при выключении эмбриональных генов. .
Незадолго до рождения происходит плавный переход от экспрессии гена -глобина плода к экспрессии гена -глобина у взрослых. Переход от фетального γ- к взрослому β-глобину не совпадает напрямую с переключением с синтеза в печени на синтез костного мозга, поскольку при рождении можно показать, что синтез как γ, так и β происходит в костном мозге.
Паттерны развития экспрессии генов α- и β-глобина.Учитывая характер активности гена глобина на протяжении внутриутробного развития и у взрослого, состав тетрамеров гемоглобина, конечно, различен.Гемоглобин плода идентифицируется как HbF и включает как α 2 Gγ 2 , так и α 2 Aγ 2 . Гемоглобин плода имеет немного более высокое сродство к кислороду, чем гемоглобин взрослого человека. Это позволяет плоду более эффективно извлекать кислород из материнского кровообращения. У взрослых основной гемоглобин идентифицируется как HbA (чаще HbA 1 ) и представляет собой тетрамер двух α и двух β цепей, как указано ранее. Минорный взрослый гемоглобин, обозначенный как HbA 2 , представляет собой тетрамер двух α-цепей и двух δ-цепей.Ген δ экспрессируется по времени, аналогичному гену β, но поскольку промотор приобрел ряд мутаций, его эффективность транскрипции снижается. Общий состав гемоглобина у здорового взрослого человека составляет приблизительно 97,5% HbA 1 , 2% HbA 2 и 0,5% HbF.
Формы гемоглобина, регулируемые развитием. Из-за паттерна развития экспрессии гена глобина различные тетрамерные формы гемоглобина присутствуют в разное время на протяжении эмбриональной, эмбриональной и взрослой жизни.Преобладающие эмбриональные тетрамеры называются Hb Gower I и Hb Gower II. Тетрамеры плода идентифицированы как Hb Portland и HbF, причем HbF является наиболее распространенной формой в развитии плода. У взрослых тетрамерный гемоглобин обозначается как HbA. HbA 1 является наиболее распространенной формой с небольшим количеством HbA 2 , образованным из белков α- и δ-глобина. У взрослых почти всегда присутствует небольшое количество HbF, которое может составлять до 0,5% от общего гемоглобина.Гемоглобинопатии
Большое количество мутаций было описано в генах глобина, включая как кластер α-глобина, так и кластер β-глобина. Эти мутации можно разделить на два разных типа: те, которые вызывают качественных аномалий (например, серповидно-клеточную анемию), и те, которые вызывают количественных аномалий (талассемии). Взятые вместе, эти расстройства называются гемоглобинопатиями . Третья группа нарушений гемоглобина включает те заболевания, при которых сохраняется экспрессия гемоглобина плода.Эти последние болезни в совокупности известны как наследственная персистенция гемоглобина плода (HPFH).
Гемоглобин S
Из мутаций, приводящих к качественным изменениям гемоглобина, наиболее распространенной является миссенс-мутация в гене β-глобина, вызывающая серповидно-клеточную анемию. Мутация, вызывающая серповидно-клеточную анемию, представляет собой замену одного нуклеотида (от A до T) в кодоне аминокислоты 6. Это изменение превращает кодон Glu (GAG) в кодон Val (GTG). Форма гемоглобина у людей с серповидно-клеточной анемией обозначается как HbS.Наследование серповидно-клеточной анемии является аутосомно-рецессивным, однако у некоторых людей при определенных условиях (например, инфекциях и лекарственной терапии) патология может наблюдаться в гетерозиготном состоянии (например, HbA / HbS).
Основная проблема серповидно-клеточной анемии заключается в том, что замена Val на Glu приводит к образованию тетрамеров гемоглобина, которые собираются в массивы при дезоксигенации в тканях. Эта агрегация приводит к деформации красных кровяных телец, делая их относительно негибкими и неспособными проходить через капиллярные русла.Повторяющиеся циклы оксигенации и деоксигенации приводят к необратимому серповидному поражению. Конечным результатом является закупорка мелких капилляров. Поскольку снижение кровотока особенно влияет на кости, возникают частые и сильные боли в костях. Это типичный симптом во время серповидно-клеточного «кризиса». Длительное периодическое закупоривание капиллярного русла приводит к повреждению внутренних органов, в частности, почек, сердца и легких. Непрерывное разрушение серповидных эритроцитов приводит к хронической анемии и эпизодам гипербилирубинемии.
Гемоглобин C
Дополнительной относительно распространенной мутацией в кодоне 6 является преобразование в кодон Lys (AAG), которое приводит к образованию HbC. Как и серповидноклеточная анемия, болезнь HbC наследуется как аутосомно-рецессивное заболевание. Люди с гомозиготным HbC / HbC проявляют относительно доброкачественное заболевание, состоящее из легкой гемолитической анемии, легкой спленомегалии и мазков крови, которые показывают присутствие кодоцитов (клеток-мишеней). Гомозиготные по HbC люди могут иметь очень легкую форму серповидноклеточной анемии.Однако у сложных гетерозигот, экспрессирующих аллель HbS и HbC (болезнь SC), существует гораздо более значимая патология.
Гемоглобин E
Гемоглобин E представляет собой форму гемоглобина, возникающую в результате мутации аминокислоты 26 в гене β-глобина. Эта мутация изменяет нормальный остаток Glu (GAG) на Lys (AAG). В дополнение к изменению аминокислотной последовательности, приводящему к качественной гемоглобинопатии, мутация HbE приводит к генерации криптического сайта сплайсинга в кодонах 25–27, что приводит к тому, что 40% мРНК β-глобина короче нормальной на 16 нуклеотидов.Эта укороченная мРНК β-глобина не дает детектируемого белка β-глобина, типичного для количественных β-талассемий.
Форма гемоглобина HbE довольно часто встречается у лиц юго-восточного азиатского происхождения. Болезнь гемоглобина Е наследуется как аутосомно-рецессивное заболевание, при котором гомозиготные люди (HbE / HbE) испытывают легкую форму β-талассемии в послеродовой период. Гетерозиготы (HbA / HbE) не проявляют никаких симптомов. Гомозиготы по аллелю HbE не проявляют симптомов при рождении из-за наличия нормального гемоглобина плода (HbF).
Симптомы начнут проявляться после прекращения экспрессии гена γ-глобина и активации гена β-глобина взрослого человека. Типичными симптомами являются легкая гемолитическая анемия и легкая спленомегалия. Серьезная патология присутствует у лиц, которые являются сложными гетерозиготами, несущими один аллель HbE и один аллель β-талассемии (HbE / β-thal). У этих людей наблюдается задержка роста, гепатоспленомегалия, гипербилирубинемия (проявляющаяся в виде желтухи), костные аномалии и сердечно-сосудистые проблемы.
Гемоглобин M
Гемоглобин M (HbM) относится к группе аутосомно-доминантных метгемоглобинемий, которые вызваны гетерозиготными мутациями в генах α- или β-глобина. Эти мутации приводят к образованию так называемого М-гемоглобина (HbM), который представляет собой форму гемоглобина, которая стабилизирует гемовое железо в состоянии трехвалентного железа (Fe 3+ ) и не поддается восстановлению. Обозначение HbM также может относиться к форме гемоглобина, которая проявляет необычную чувствительность к окислителям.
Было охарактеризовано по крайней мере пять различных форм гемоглобина М. Четыре из них, HbM (Бостон), HbM (Гайд-Парк), HbM (Иватэ) и HbM (Саскатун), обусловлены мутациями, которые изменяют критические остатки His на Tyr в гемосвязывающем кармане α-глобина или β- глобиновые белки. Пятая форма HbM (Milwaukee-1) является результатом замены Glu на Val в положении четырех аминокислотных остатков от дистального His гема-связывающего кармана. Все эти мутации стабилизируют гемовое железо в трехвалентном (окисленном) состоянии.
Основной патологией у пациентов с мутациями HbM является цианоз, но в остальном они протекают бессимптомно. Если мутация HbM присутствует в гене α-глобина, цианоз проявляется при рождении. Если мутация HbM находится в гене β-глобина, то цианоз появляется позже или усиливается при увеличении продукции β-субъединицы. Новорожденные, у которых есть мутации в гене γ-глобина, будут демонстрировать цианоз при рождении, но он исчезнет, когда произойдет полное переключение экспрессии гена γ-глобина на β-глобин.
Электрофорез гемоглобина
Электрофорез белков гемоглобина от людей с подозрением на серповидно-клеточную анемию (или несколько других типов нарушений гемоглобина) является эффективным диагностическим инструментом, потому что варианты гемоглобинов имеют разные заряды и, следовательно, по-разному мигрируют при гель-электрофорезе. Пример этого метода показан на рисунке ниже.
Образец электрофореза гемоглобина у нескольких разных людей. Дорожки 1 и 5 представляют собой стандарты гемоглобина.Дорожка 2 — нормальный взрослый. Дорожка 3 — нормальный новорожденный. Дорожка 4 представляет собой гомозиготную особь по HbS. Дорожки 6 и 8 представляют собой гетерозиготные HbA / HbS индивидуумы. Дорожка 7 представляет собой индивидуум, гетерозиготный по соединению HbS / HbC.Еще одним эффективным инструментом для определения генотипа лиц с подозрением на серповидноклеточную анемию, а также для пренатальной диагностики является выполнение ПДРФ-картирования или использование ПЦР. Пример использования этих инструментов можно увидеть на странице «Молекулярные инструменты медицины».
В дополнение к миссенс-мутациям, которые приводят к HbS, HbC, HbE и HbM, был идентифицирован ряд мутаций сдвига рамки считывания, приводящих к качественным отклонениям в гемоглобине.Вставка 2 нуклеотидов между кодонами 144 и 145 в гене β-глобина приводит к образованию гемоглобина Cranston . Вставка, которая находится рядом с С-концом белка β-глобина, приводит к тому, что нормальный стоп-кодон выходит за пределы рамки и синтез продолжается в 3′-нетранслируемую область до случайного стоп-кодона. В результате получается белок β-глобин из 157 аминокислот.
В варианте с константой гемоглобина мутация в гене α-глобина превращает стоп-кодон (UAA) в кодон глутамина (Gln) (CAA), так что длина белка оказывается на 31 аминокислоту длиннее, чем обычно.Образующийся альфа-глобиновый белок в гемоглобине Constant Spring не только качественно изменен, но и потому, что он нестабилен, это также количественное отклонение.
Поскольку локус глобинового гена содержит кластеры схожих генов, существует возможность неравного кроссовера между сестринскими хроматидами во время профазы мейоза I. Генерация гемоглобина Gun Hill и гемоглобина Lepore является результатом неравного скрещивания. над событиями. Гемоглобин Gun Hill является результатом делеции 15 нуклеотидов, вызванной неравным кроссинговером между кодонами 91–94 одного гена β-глобина и кодонами 96–98 другого.Генерация гемоглобинов Lepore является результатом неравного кроссовера между генами δ-глобина и β-глобина. Получающийся в результате гибридный ген δβ называется Lepore, а гибридный ген βδ называется анти-Lepore. Как указывалось ранее, промотор гена δ-глобина неэффективен, поэтому последствия этого неравномерного кроссовера являются как качественными, так и количественными.
Талассемия
Талассемии представляют собой количественные гемоглобинопатии, которые возникают в результате нарушений синтеза гемоглобина и влияют на кластеры генов как α-глобина, так и β-глобина.Недостаток синтеза β-глобина приводит к β-талассемии, а недостаток синтеза α-глобина приводит к α-талассемии. Термин талассемия происходит от греческого слова thalassa, означающего «море», и применялся к этим расстройствам из-за высокой частоты их возникновения у людей, живущих вокруг Средиземного моря.
У нормальных людей образуется равное количество белков α- и β-глобина, что позволяет им стехиометрически объединяться с образованием правильных тетрамеров гемоглобина.При α-талассемии образуется нормальное количество β-глобина, но не белков α-глобина. Белки β-глобина способны образовывать гомотетрамеры (β 4 ), и эти тетрамеры называются гемоглобином H (HbH). Избыток HbH в эритроцитах приводит к образованию телец включения, обычно наблюдаемых у пациентов с α-талассемией. Кроме того, тетрамеры HbH обладают заметно сниженной кислородной емкостью. При β-талассемии, где β-глобины недостаточны, α-глобины находятся в избытке и образуют гомотетрамеры α-глобина.Гомотетрамеры альфа-глобина крайне нерастворимы, что приводит к преждевременному разрушению эритроцитов в костном мозге и селезенке.
При α-талассемии уровень продукции α-глобина может варьироваться от нулевого до почти нормального. Частично это связано с тем, что на хромосоме 16 есть два идентичных гена альфа-глобина. Таким образом, альфа-талассемия включает инактивацию от одного до всех четырех генов альфа-глобина. Если три из четырех генов альфа-глобина функционируют, люди полностью бессимптомны.Эта ситуация определяется как состояние «молчащего носителя» или иногда как α-талассемия 2. Генотипически эта ситуация обозначается как αα / α– (где прочерк указывает на нефункциональный ген) или α– / αα. Если два из четырех генов инактивированы, индивидов обозначают как признак α-талассемии или как α-талассемия 1. Генотипически эта ситуация обозначается как αα / — -. У лиц африканского происхождения с α-талассемией 1 заболевание обычно возникает в результате инактивации одного гена α-глобина на каждой хромосоме и обозначается α– / α–.Это означает, что эти люди гомозиготны по хромосоме α-талассемии 2. Фенотип α-талассемии 1 относительно доброкачественный. Средний объем эритроцитов (в клинических испытаниях обозначенный как MCV) снижается при α-талассемии 1, но у людей обычно нет симптомов.
Клиническая ситуация становится более тяжелой, если функционирует только один из четырех генов α-глобина. Из-за резкого снижения продукции цепи α-глобина в этой последней ситуации присутствует высокий уровень тетрамера β 4 .Клинически это заболевание обозначается как гемоглобин H (HbH) . Больные имеют анемию от умеренной до выраженной, и их MCV довольно низкий, но болезнь не смертельна.
Наиболее тяжелая ситуация возникает, когда не образуются цепи α-глобина (генотипически обозначенные — — / — -). Это приводит к внутриутробной летальности или ранней неонатальной смерти. Преобладающий гемоглобин плода у пораженных людей представляет собой тетрамер γ-цепей и обозначается как гемоглобин Barts .Этот гемоглобин имеет очень высокое сродство к кислороду, что приводит к плохому выделению кислорода и, следовательно, к кислородному голоданию в тканях плода. Сердечная недостаточность возникает из-за того, что сердце пытается перекачивать больше насыщенной кислородом крови к тканям, страдающим кислородным голоданием, что приводит к заметному отеку. Последняя ситуация называется водянкой плода .
Было идентифицировано большое количество мутаций, приводящих к уменьшению или отсутствию продукции цепей β-глобина, что приводит к β-талассемии. В наиболее тяжелой ситуации мутации как материнского, так и отцовского генов β-глобина приводят к потере нормального количества белка β-глобина.Полное отсутствие HbA обозначается как β 0 -талассемия. Если та или иная мутация позволяет продуцировать небольшое количество функционального β-глобина, то нарушение обозначается как β + -талассемия.
Обе талассемии β 0 и β + обозначаются как большая талассемия , также называемая анемией Кули в честь доктора Томаса Кули, который первым описал это заболевание. Больные страдают тяжелой анемией, начиная с первого года жизни, что приводит к необходимости переливания крови.Вследствие анемии костный мозг резко увеличивает свои усилия по производству крови. Кора кости истончается, что приводит к патологическому перелому и деформации костей лица и черепа. Кроме того, имеется выраженная гепатоспленомегалия, поскольку печень и селезенка выступают в качестве дополнительных участков кроветворения. Без вмешательства эти люди умрут в течение десятилетия жизни. Как указано, пациенту с большой β-талассемией требуются переливания крови, однако в долгосрочной перспективе эти переливания приводят к накоплению железа в органах, особенно в сердце, печени и поджелудочной железе.Отказ органа приводит к смерти в подростковом возрасте до 20 лет. Терапия хелатированием железа, по-видимому, улучшает прогноз для пациентов с большой β-талассемией, но для этого требуется постоянная инфузия хелатирующего агента.
Лица, гетерозиготные по β-талассемии, имеют так называемую малую талассемию . Больные люди несут в себе один нормальный ген β-глобина и другой ген, который несет мутацию, приводящую к уменьшению выработки β-глобина или его отсутствию. Лица, которые не производят какой-либо функциональный белок -глобин из одного гена, называются гетерозиготами β 0 .Если продукция β-глобина снижена в одном локусе, особей называют гетерозиготами β + . У лиц с малой талассемией симптомы обычно протекают бессимптомно.
Термин промежуточная талассемия используется для обозначения лиц со значительной анемией, у которых имеются симптомы, но, в отличие от большой талассемии, переливания крови не требуются. Этот синдром приводит к индивидуумам, у которых оба гена β-глобина экспрессируют пониженное количество белка или когда один ген не производит его, а другой дает умеренно пониженное количество.Человек, который является сложной гетерозиготой с α-талассемией и β + -талассемией, также будет проявляться как промежуточная талассемия.
Основной причиной α-талассемии является делеция, тогда как для β-талассемии мутации более тонкие. При β-талассемиях были охарактеризованы точечные мутации в промоторе, мутации в кодоне инициации трансляции, точечные мутации в сигнале полиаденилирования и множество мутаций, ведущих к аномалиям сплайсинга.
Наследственная устойчивость гемоглобина плода: HPFH
Есть люди, у которых время развития продукции глобина изменяется в результате мутации.Люди с наследственной персистентностью фетального гемоглобина (HPFH) продолжают вырабатывать фетальный гемоглобин (HbF) во взрослом возрасте. В то время как люди с γβ-талассемией имеют умеренно повышенные уровни HbF и умеренный, но определенный фенотип талассемии, состоящий из гипохромии и микроцитоза, HPFH обычно не связан с гипохромией и микроцитозом у гетерозигот. Поскольку синдром HPFH является доброкачественным, большинство людей даже не подозревают, что они являются носителями аномального гемоглобина. Многие индивидуумы HPFH несут делеции в кодирующих областях гена δ-глобина и / или β-глобина в кластере генов β-глобина.Кроме того, некоторые индивидуумы HPFH обладают мутациями в промоторной области генов γ-глобина плода. У лиц, гомозиготных по мутациям HPFH, отсутствуют все формы гемоглобина взрослого человека HbA 1 и HbA 2 .
Шесть различных делеций были идентифицированы в кластере генов β-глобина, которые связаны с фенотипом HPFH. Делеции HPFH-1 и HPFH-2 представляют собой большие делеции в генах δ- и β-глобина. Делеции HPFH-1 и HPFH-2 имеют почти одинаковый размер и охватывают приблизительно 105 т.п.н. ДНК.Делеции HPFH-3 и HPFH-4 менее обширны. Эти две делеции охватывают приблизительно 50 т.п.н. и 40 т.п.н., разделенные примерно 2 т.п.н., и они расположены примерно на 30 т.п.н. ниже положения гена β-глобина. Делеция HPFH-5 представляет собой относительно короткую делецию, которая простирается от точки примерно 3 т.п.н. 5 ‘гена δ-глобина до точки 3’ гена β-глобина 700 п.н. Делеция HPFH-5 проксимальна (3 ‘) по отношению к положению 3′ энхансера гена β-глобина. Делеция HPFH-6 составляет приблизительно 101 т.п.н., но больше в 5’-направлении и связана с делецией гена Aγ-глобина плода.
Ряд различных точечных мутаций в промоторных областях генов гамма-глобина плода был идентифицирован у индивидуумов HPFH. Большинство этих промоторных мутаций затрагивают ген Aγ-глобина. В любом случае результатом является сверхэкспрессия генов гамма-глобина плода. Было обнаружено, что эти точечные мутации сгруппированы в трех различных областях 5′-фланкирующей ДНК затронутых генов γ-глобина. В одной из этих областей было охарактеризовано пять различных мутаций, все в области на 200 п.н. выше сайта начала транскрипции генов γ-глобина.Четыре из этих мутаций находятся в промоторной области гена Aγ-глобина и одна — в области промотора гена Gγ-глобина. Все эти мутации находятся в GC-богатых областях этих двух генов, которые, как известно, связывают фактор транскрипции Sp1.
В дополнение к связыванию Sp1, эта область генов γ-глобина также связывает другой комплекс регуляции транскрипции, который специфичен для эритроидных клеток. Область мутации второй области промотора расположена в положении -175. Точечная мутация, изменяющая T на C, обнаруживается в этом месте в любом из генов γ-глобина.Эта область обоих генов содержит сайты связывания для факторов транскрипции OCT-1 и GATA-1. Точечная мутация в положении -175 затрагивает один нуклеотид, который присутствует в частично перекрывающихся сайтах связывания для обоих этих факторов транскрипции. Третья область, затронутая точечной мутацией, находится в области CCAAT-бокса гена Aγ-глобина. Другая мутация в гене Aγ-глобина, приводящая к подобному фенотипу, включает делецию 13 пар оснований ДНК с положения -102 до -114, которая охватывает бокс-элемент CCAAT.Также была идентифицирована третья мутация, которая включает CCAAT-бокс гена Gγ-глобина.
Анализ алгоритмов | Набор 3 (асимптотические обозначения)
Мы обсудили асимптотический анализ, наихудшие, средние и наилучшие варианты алгоритмов. Основная идея асимптотического анализа состоит в том, чтобы иметь меру эффективности алгоритмов, которая не зависит от констант, зависящих от машины, и не требует реализации алгоритмов и времени, затрачиваемого программами на сравнение. Асимптотические обозначения — это математические инструменты для представления временной сложности алгоритмов асимптотического анализа.Следующие 3 асимптотические обозначения в основном используются для представления временной сложности алгоритмов.
1) Θ Обозначение: Обозначение тета ограничивает функции сверху и снизу, поэтому оно определяет точное асимптотическое поведение.
Простой способ получить тета-нотацию выражения — отбросить младшие члены и игнорировать ведущие константы. Например, рассмотрим следующее выражение.
3n 3 + 6n 2 + 6000 = Θ (n 3 )
Удаление членов более низкого порядка всегда нормально, потому что всегда будет n0, после которого Θ (n 3 ) будет иметь более высокие значения, чем Θn 2 ) независимо от используемых констант.
Для данной функции g (n) обозначим Θ (g (n)) — это следующий набор функций.
Θ (g (n)) = {f (n): существуют положительные постоянные c1, c2 и n0 такие
что 0 <= c1 * g (n) <= f (n) <= c2 * g (n) для всех n> = n0} Приведенное выше определение означает, что если f (n) является тэтой g (n), то значение f (n) всегда находится между c1 * g (n) и c2 * g (n) для больших значений n (n> = n0). Определение теты также требует, чтобы f (n) была неотрицательной для значений n больше n0.2).
2. Наилучшая временная сложность сортировки вставкой равна Θ (n).
Нотация Big O полезна, когда у нас есть только верхняя граница временной сложности алгоритма. Часто мы легко находим верхнюю границу, просто глядя на алгоритм.
O (g (n)) = {f (n): существуют положительные постоянные c и
n0 такое, что 0 <= f (n) <= c * g (n) для
все n> = n0} 3) Ω Обозначение: Подобно тому, как обозначение Big O обеспечивает асимптотическую верхнюю границу функции, обозначение Ω обеспечивает асимптотическую нижнюю границу.
Ω Обозначение может быть полезно, когда у нас есть нижняя граница временной сложности алгоритма. Как обсуждалось в предыдущем посте, наилучшая производительность алгоритма обычно бесполезна, нотация Омега — наименее используемая нотация среди всех трех.
Для заданной функции g (n) обозначим через Ω (g (n)) множество функций.
Ω (g (n)) = {f (n): существуют положительные постоянные c и
n0 такое, что 0 <= c * g (n) <= f (n) для
все n> = n0}. Давайте рассмотрим здесь тот же пример сортировки вставкой. Временную сложность сортировки вставкой можно записать как Ω (n), но это не очень полезная информация о сортировке вставкой, поскольку нас обычно интересуют наихудший случай, а иногда и средний случай.
Свойства асимптотических обозначений:
После того, как мы прошли определение этих трех обозначений, давайте теперь обсудим некоторые важные свойства этих обозначений.
- Общие свойства:
Если f (n) равно O (g (n)), то a * f (n) также O (g (n)); где а — постоянная.
Пример: f (n) = 2n² + 5 равно O (n²)
, тогда 7 * f (n) = 7 (2n² + 5)
= 14n² + 35 также O (n²)Аналогичным образом это свойство удовлетворяется как для обозначений, так и для Ω.
Можно сказать
Если f (n) равно Θ (g (n)), то a * f (n) также равно Θ (g (n)); где а — постоянная.
Если f (n) является Ω (g (n)), то a * f (n) также является Ω (g (n)); где а — постоянная. - Отражающие свойства:
Если задано f (n), то f (n) равно O (f (n)).
Пример: f (n) = n²; O (n²) я.e O (f (n))
Точно так же это свойство удовлетворяется как для обозначений, так и для Ω.
Можно сказать
Если задано f (n), то f (n) равно Θ (f (n)).
Если задано f (n), то f (n) равно Ω (f (n)). - Переходные свойства:
Если f (n) равно O (g (n)) и g (n) равно O (h (n)), то f (n) = O (h (n)).
Пример: если f (n) = n, g (n) = n² и h (n) = n³
n равно O (n²), а n² равно O (n³)
, то n равно O (n³)Аналогичным образом это свойство удовлетворяется как для обозначений, так и для Ω.
Можно сказать
Если f (n) равно Θ (g (n)) и g (n) равно Θ (h (n)), то f (n) = Θ (h (n)).
Если f (n) равно Ω (g (n)) и g (n) равно Ω (h (n)), то f (n) = Ω (h (n)) - Симметричные свойства:
Если f (n) равно Θ (g (n)), то g (n) равно Θ (f (n)).
Пример: f (n) = n² и g (n) = n²
, тогда f (n) = Θ (n²) и g (n) = Θ (n²)Это свойство удовлетворяет только обозначению Θ.
- Симметричные свойства транспонирования:
Если f (n) равно O (g (n)), то g (n) равно Ω (f (n)).
Пример: f (n) = n, g (n) = n²
, тогда n равно O (n²), а n² равно Ω (n)Это свойство выполняется только для обозначений O и Ω .
- Еще свойства:
- Если f (n) = O (g (n)) и f (n) = Ω (g (n)), то f (n) = Θ (g (n))
- Если f (n) = O (g (n)) и d (n) = O (e (n))
, то f (n) + d (n) = O (max (g (n), e ( n)))
Пример: f (n) = n, т.е. O (n)
d (n) = n², т.е. O (n²)
, тогда f (n) + d (n) = n + n², т.е. O (n²) - Если f (n) = O (g (n)) и d (n) = O (e (n))
, то f (n) * d (n) = O (g (n) * e (n) )
Пример: f (n) = n i.2)
3. Для любых двух функций f (n) и g (n) имеем f (n) = Θ (g (n)) тогда и только тогда, когда f (n) = O (g (n) ) и f (n) = Ω (g (n)).
4. Временная сложность всех компьютерных алгоритмов может быть записана как Ω (1) Важные ссылки:Ссылки:
Lec 1 | MIT (Введение в алгоритмы)Автор статьи: Abhay Rathi . Пожалуйста, напишите комментарии, если вы обнаружите что-то неправильное, или вы хотите поделиться дополнительной информацией по теме, обсужденной выше.
Вниманию читателя! Не прекращайте учиться сейчас. Освойте все важные концепции DSA с помощью курса DSA Self Paced Course по доступной для студентов цене и подготовьтесь к работе в отрасли.
.
