Анатомия нижней конечности
Области нижней конечности : Иллюстрации: А. Мишо , E-anatomy, Imaios
Кости верхней конечности , Соединения нижней конечности : Рисунки
Тазовая кость , Вид сбоку : Трехмерные изображения
Тыл стопы , Анатомия : Кости; система скелета, Суставы стопы
Коленный сустав , Большеберцовая кость/Верхняя суставная поверхность : Вид сверху
Мышцы; мышечная система , Человеческое тело : Переднее фасциальное ложе бедра; фасциальное ложе разгибателей
Предплюсна : Медиальный вид
Артерии нижней конечности , Бедро : Вид спереди
Срез , Анатомия (Поперечный/Аксиальный): Тазобедренный сустав, Ягодицы, Тазовый пояс; пояс нижней конечности, Бедро
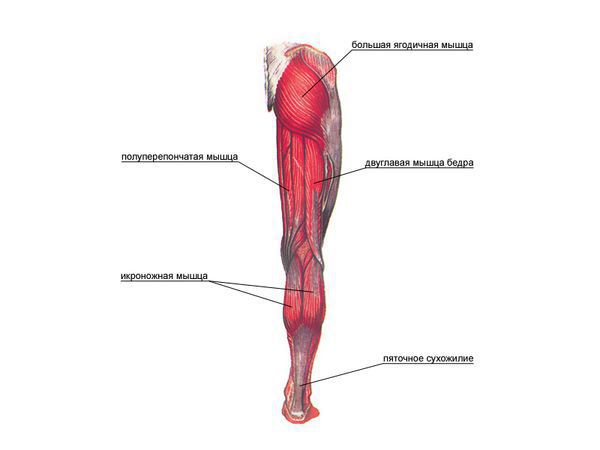
Срез , Человеческое тело (Аксиальный/Поперечный) : Голень/Икра
Лечение варикоза в Челябинске | лечимварикоз.
 рф
рфРис. 1 Правильная работа венозных клапанов
В нашем теле теле есть два типа кровеносных сосудов – артерии и вены. С помощью артерий богатая кислородом кровь от легких и сердца поступает ко всем органам и тканям, в том числе и к ногам. Функция вен – отводить бедную кислородом кровь обратно к сердцу и легким. Для того чтобы кровь от ног бежала вверх против силы тяжести, существуют специальные венозные клапаны, которые пропускают ее только в одном направлении (Рисунок 1). Во время ходьбы происходит сокращение мышц голени, они сжимают глубокие вены, и кровь выбрасывается вверх. Этот механизм называется венозно-мышечная помпа. Именно поэтому пациентам с варикозом рекомендуют больше лежать или ходить, и меньше стоять или сидеть.
В ногах выделяют глубокие и подкожные (поверхностные) вены, а также соединяющие эти две системы между собой, так называемые перфоранты. При растяжении вен створки клапанов перестают дотягиваться друг до друга, и кровь протекает между ними обратно.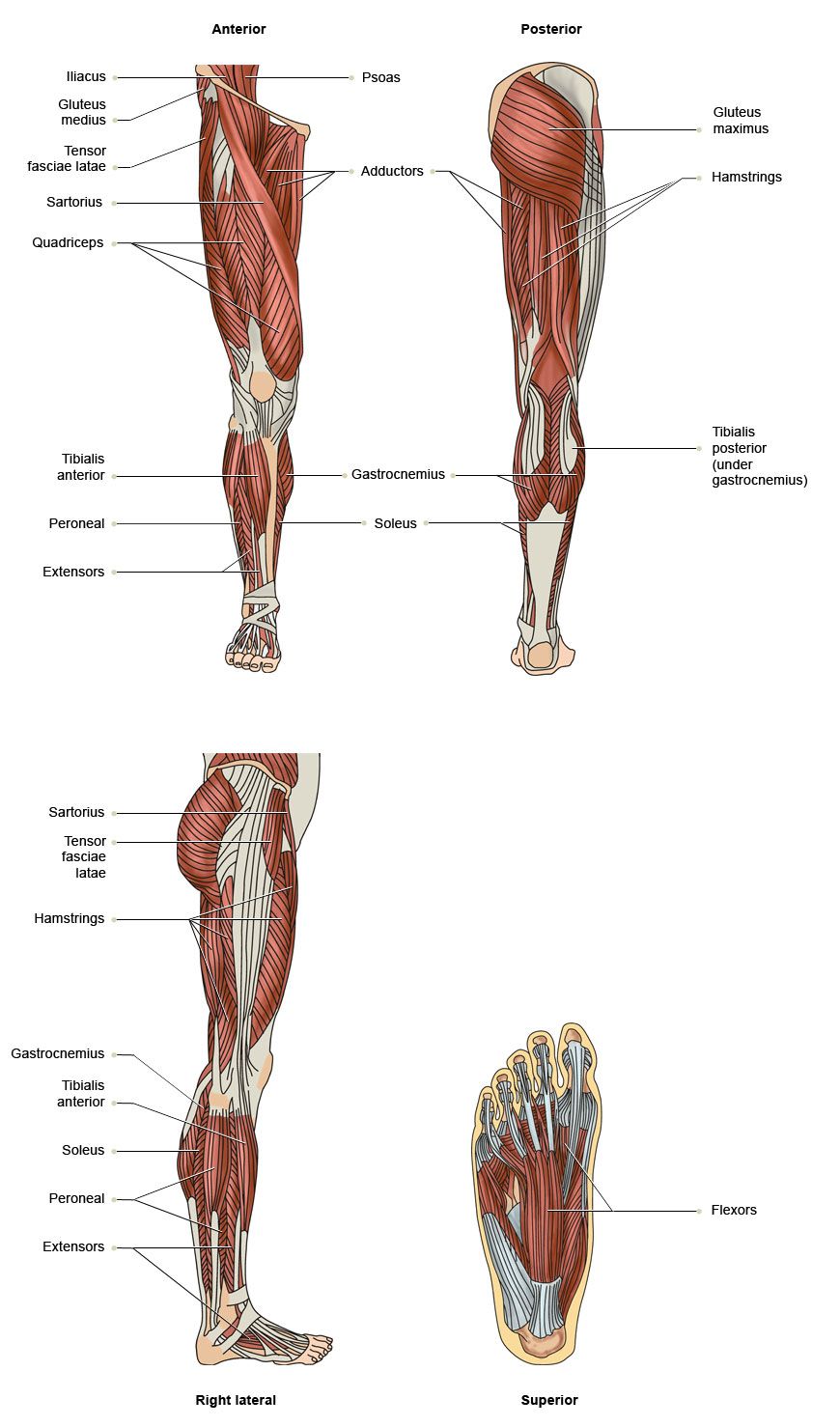 Это и называется варикозным расширением. Подробнее об этом можно прочитать здесь.
Это и называется варикозным расширением. Подробнее об этом можно прочитать здесь.
Нужно отметить, что в подавляющем большинстве случаев варикозному расширению подвергаются не глубокие, а именно поверхностные вены, окруженные снаружи мягкой подкожно-жировой клетчаткой, а не плотными мышцами, костями и связками.
Часто люди приходят с убеждением, что у них “варикоз глубоких вен”. В таких случаях, либо у пациента есть боли в ногах, причину которых никто не смог установить, и в итоге эту боль списали на “мифический диагноз”, либо речь идет о поражении магистральных подкожных вен, которые располагаются глубже, чем их притоки, но все равно относятся именно к подкожной (поверхностной) венозной системе.
Подкожные вены представлены двумя основными стволами – большой и малой подкожными венами (Рисунок 2). У худых людей в вертикальном положении достаточно часто их можно увидеть невооруженным глазом. Большая подкожная вена идет от внутренней лодыжки вверх по внутренней поверхности голени и бедра и в области паховой складки впадает в глубокую венозную систему. Анатомия малой подкожной вены достаточно разнообразна, но в большинстве случаев она поднимается от наружной лодыжки по задней поверхности голени и впадает в глубокие вены в подколенной области.
Анатомия малой подкожной вены достаточно разнообразна, но в большинстве случаев она поднимается от наружной лодыжки по задней поверхности голени и впадает в глубокие вены в подколенной области.
Рис. 2 Схематическая анатомия подкожных вен нижних конечностей
Нужно сказать, что у большинства пациентов с варикозом вен нижних конечностей мы видим не магистральные подкожные вены, а их притоки, то есть те вены, которые в них впадают (рисунок 3).
Бывает также расширение самых маленьких, внутрикожных вен, которые еще называют “сосудистыми звездочками” (Рисунок 4). Это отдельное заболевание, о котором мы рассказываем в соответствующем разделе сайта.
Рис. 3 Расширенные притоки большой подкожной вены (саму большую подкожную вену (ствол) не видно, видно только ее притоки – ветки)
Рис. 4 Сосудистые звездочки (расширенные внутрикожные вены)
Все методики лечения варикоза направлены на то, чтобы устранить расширенные подкожные вены. Самый частый вопрос пациентов при этом: “А как же тогда кровь будет бежать обратно?” Но, как мы сказали ранее, по варикозным венам кровь уже не просто не бежит вверх в сторону сердца, а даже наоборот – бедная кислородом кровь между клапанами протекает вниз. То есть пациенты уже живут не просто без этих вен, а еще и в условиях того, что эти вены вредят. Таким образом, на здоровые вены ложится повышенная нагрузка, а когда мы убираем варикозные, расширенные вены, здоровым от этого становится только легче. Кроме того, у Вас всегда есть глубокая венозная система, которая, как было сказано выше, практически никогда не подвергается варикозному расширению, так как снаружи окружена плотными мышцами, костями и связками, а не мягкой жировой клетчаткой.
То есть пациенты уже живут не просто без этих вен, а еще и в условиях того, что эти вены вредят. Таким образом, на здоровые вены ложится повышенная нагрузка, а когда мы убираем варикозные, расширенные вены, здоровым от этого становится только легче. Кроме того, у Вас всегда есть глубокая венозная система, которая, как было сказано выше, практически никогда не подвергается варикозному расширению, так как снаружи окружена плотными мышцами, костями и связками, а не мягкой жировой клетчаткой.
На сегодняшний день самым современным методом лечения магистральных подкожных вен является эндовенозная лазерная коагуляция, а их притоков – минифлебэктомия и склеротерапия.
Тайна эволюции: откуда ноги растут…
Подпись к фото,Исследователи снимали мерку с конечностей человека и шимпанзе
Ученые, возможно, раскрыли тайну эволюции человеческих рук, в свое время развившихся настолько, чтобы изготовить и использовать каменные орудия труда.
Группа ученых на страницах журнала Evolution сообщила, что изменения наших рук и пальцев на них стали побочным эффектом изменения формы человеческих ступней.
Все это, по их словам, говорит о том, что появление каменных орудий труда было напрямую связано со способностью человека стоять и передвигаться на двух конечностях.
Ученые использовали математическую модель для того, чтобы воспроизвести изменения, происходившие с конечностями человека.
«Это возвращает нас к дарвиновскому «Происхождению человека», — полагает руководитель проекта Кэмпбелл Рольен из университета Калгари в Канаде.
По его словам, именно Дарвин одним из первых предположил, что существует связь между технологией производства каменных орудий труда и двуногим хождением.
«Его идея состояла в том, что это были два разных события, произошедшие одно за другим, и что хождение на двух ногах освободило руки для других целей», — заявил Рольен.
Параллельное развитие
В ходе исследования доктор Рольен с коллегами снимали мерку с ладоней и ступней людей и шимпанзе.
Результаты измерений показали сильную взаимосвязь между аналогичными частями ладоней и ступней.
«Если у вас длинный большой палец руки, то скорее всего, большой палец ноги тоже будет длинней других», — пояснил доктор Рольен.
«Одну из причин сильной корреляции между пальцами рук и ног мы видим в том, что их развитие происходит на основе одного и того же генетического рисунка, и любое незначительное изменение этого рисунка может параллельно отразиться на руках и ногах», — сказал исследователь.
Затем на основе собранной базы данных ученые приступили к созданию математической модели, симулирующей эволюционные изменения конечностей.
Модель, в числе прочего, воссоздавала мельчайшие подробности изменений, чтобы отследить производимый ими эффект.
Почему шимпанзе?
На основе математической модели исследователи выяснили, что изменения формы ступней вызывали параллельные изменения формы ладоней.
Предположительно, это привело к тому, что наши предки, включая неандертальцев, сумели развить ловкость рук, необходимую для создания каменных орудий труда.
Робин Кромптон, профессор анатомии Ливерпульского университета, назвал исследование канадских коллег интересным, однако, по его мнению, оно вызывает ряд вопросов.
«Лично мне не кажется, что руки и ноги шимпанзе являются хорошей моделью рук и ног предков человека. В этой связи более подходящими были бы руки и ноги равнинной гориллы», — заявил Кромптон в интервью Би-би-си.
Кроме того, по мнению профессора, необходимо анализировать не только и не столько размеры конечностей человека, сколько функциональную форму и биомеханику.
Его коллега, профессор Пол О’Хиггинс из Йоркской медицинской школы города Халла, высказал мнение, что результаты исследования канадских ученых получат дальнейшее развитие и могут быть дополнены новыми данными.
Стопа человека особенности строения | Ortofoot
Стопа человека анатомия и физиология
Функции стопы
Стопа человека несет на себе вес всего тела.
Анатомия стопы
Каждая стопа человека состоит из 26 костей. Кости стопы широкие и плоские и связаны между собой большим количеством прочных связок, которые ограничивают движения, но усиливают стопу как опору. Прочность стопы как целой конструкции важна при совершении движений тела и удержании его веса. Несмотря на ограниченную подвижность. стопа может легко перемещаться как по гладкой, так и по не ровной поверхности.
Стопа разделяется на 3 части: 1.Кости пальцев стопы. 2.Кости плюсны. 3.Кости предплюсны.
Каждый палец стопы (всего их пять) имеет 3 фаланги, за исключением большого пальца стопы, который имеет 2 фаланги. Кости пальцев стопы соединяются с костями плюсны.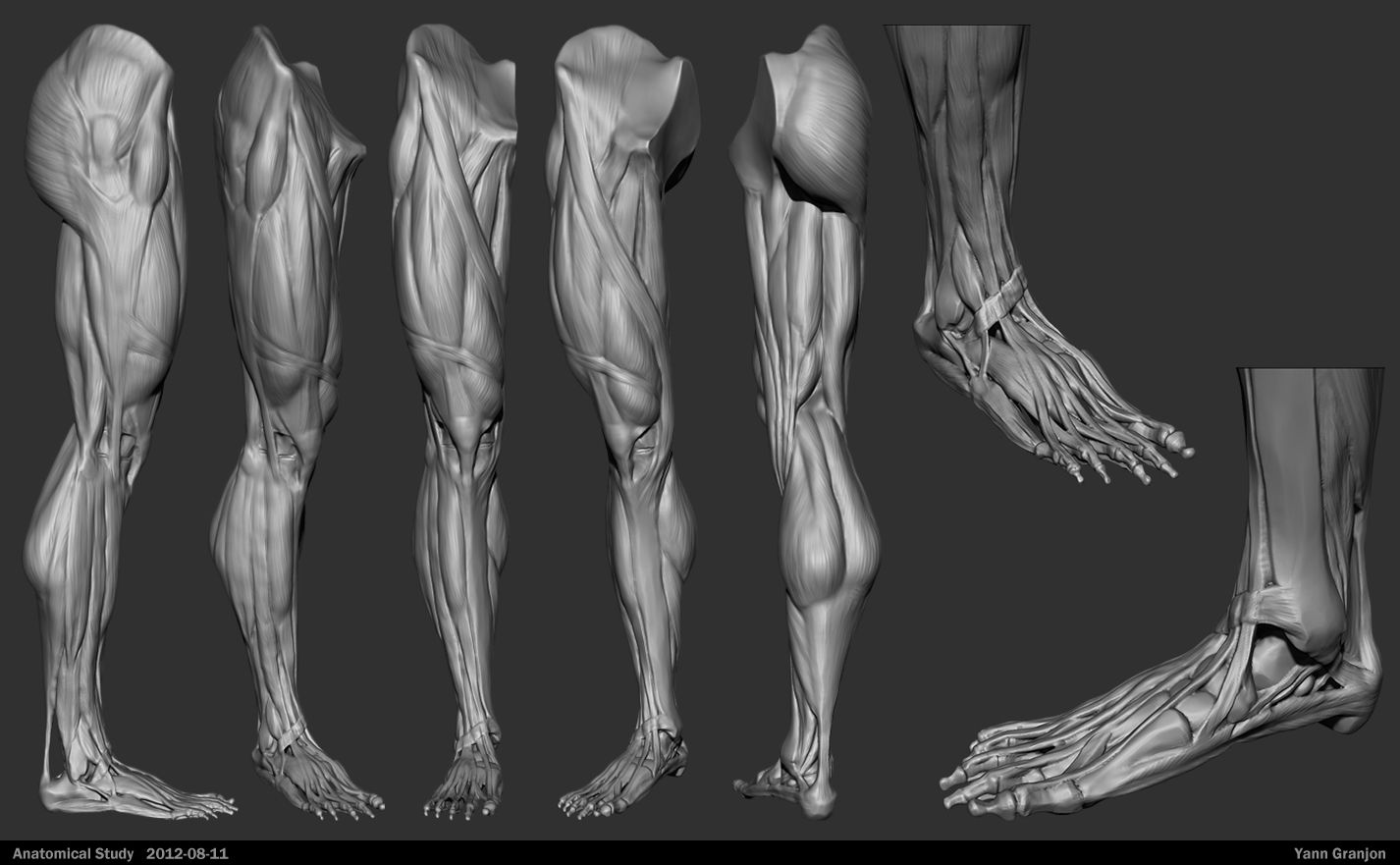
 Анатомия костей стопы.
Анатомия костей стопы.В стопе много сложных суставов. Пяточная кость вместе с таранной костью сзади и кубовидная с ладьевидной костью спереди образуют, так называемый по автору, комбинированный сустав Шопара. Кубовидная кость и три клиновидные кости сзади, а также пять плюсневых костей спереди образуют сустав Лисфранка. Сустав Шопара называют еще поперечным суставом предплюсны. Кости предплюсны и плюсны, а также связывающие их сухожилия и связки образуют арки стопы или своды, которые поднимают стопу над поверхностью. Аркообразные своды стопы за счет амортизации гасят нарузки, возникающие при ходьбе и беге. Сначала стопа человека уплощается, а затем вновь принимает выгнутую форму. Также арки, образованные костями предплюсны и плюсны, связки, соединяющие их действуют как подъемный механизм, толкающий тело вверх при ходьбе и беге.
Своды стопы.
В стопе различают пять продольных сводов и один поперечный свод стопы.Продольные своды стопы начинаются от пятки и продолжаются по выпуклым линиям к плюсневым костям стопы.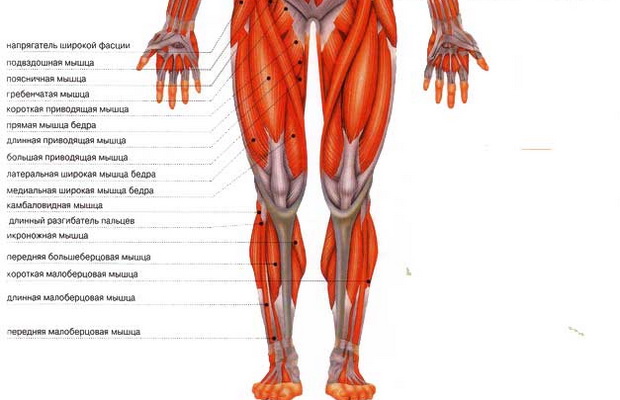
Своды стопы формируют как кости стопы, так и сухожилия и связки, и мышцы. Продольные мышцы стопы укорачивают и увеличивают продольные своды, а косые мышцы сужают стопу и увеличивают поперечный свод. Самая мощная связка, формирующая и удерживающая продольный свод — длинная подошвенная связка.
Стопа человека, вид сбоку (медиальная сторона).
Когда эта конструкция ослабевает происходит проседание сводов и стопа уплощается, иногда это сочетается с деформацией суставов стопы.
Длинные, сильные и широкие кости ноги и стопы обеспечивают устойчивость тела, удерживают его вес и устойчивость тела, удерживают его вес и распределяют силу, генерируемую при беге и прыжках.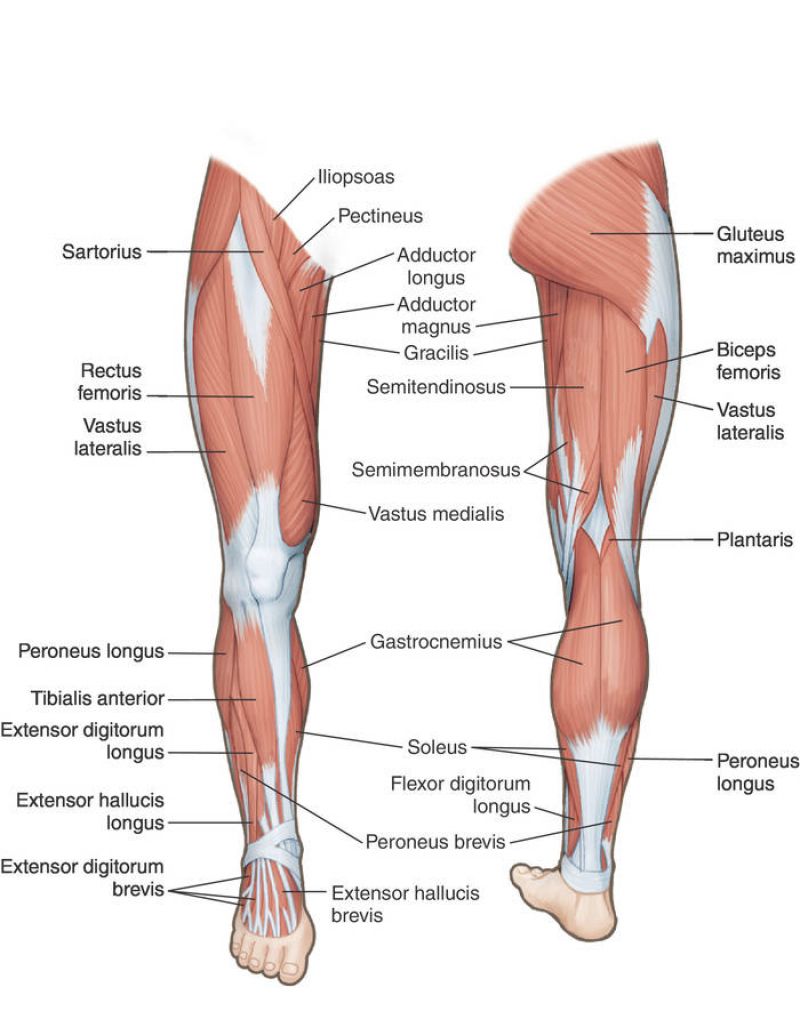 Каждая нижняя конечность состоит из трех частей: бедра, голени и стопы. (Количество костей нижних конечностей — 30).
Каждая нижняя конечность состоит из трех частей: бедра, голени и стопы. (Количество костей нижних конечностей — 30).
Венозная система ног | Venenzentrum am See
Анатомия венозной системы нижних конечностей отличается большой вариабельностью. Большую роль в оценке данных инструментального обследования в выборе правильного метода лечения играет знание индивидуальных особенностей строения венозной системы человека.Глубокая венозная сеть: Передние и задние большеберцовые вены сливаются в бедренно-подколенном канале и образуют непарную подколенную вену, которая переходит в мощный ствол бедренной вены (v. femoralis). В бедренную вену, еще до перехода в наружную подвздошную (v. iliaca externa), впадают 5-8 перфорантных вен и глубокая вена бедра (v. femoralis profunda), несущую кровь от мышц задней поверхности бедра.
Поверхностная венозная сеть: Расположена в подкожной клетчатке над поверхностной фасцией.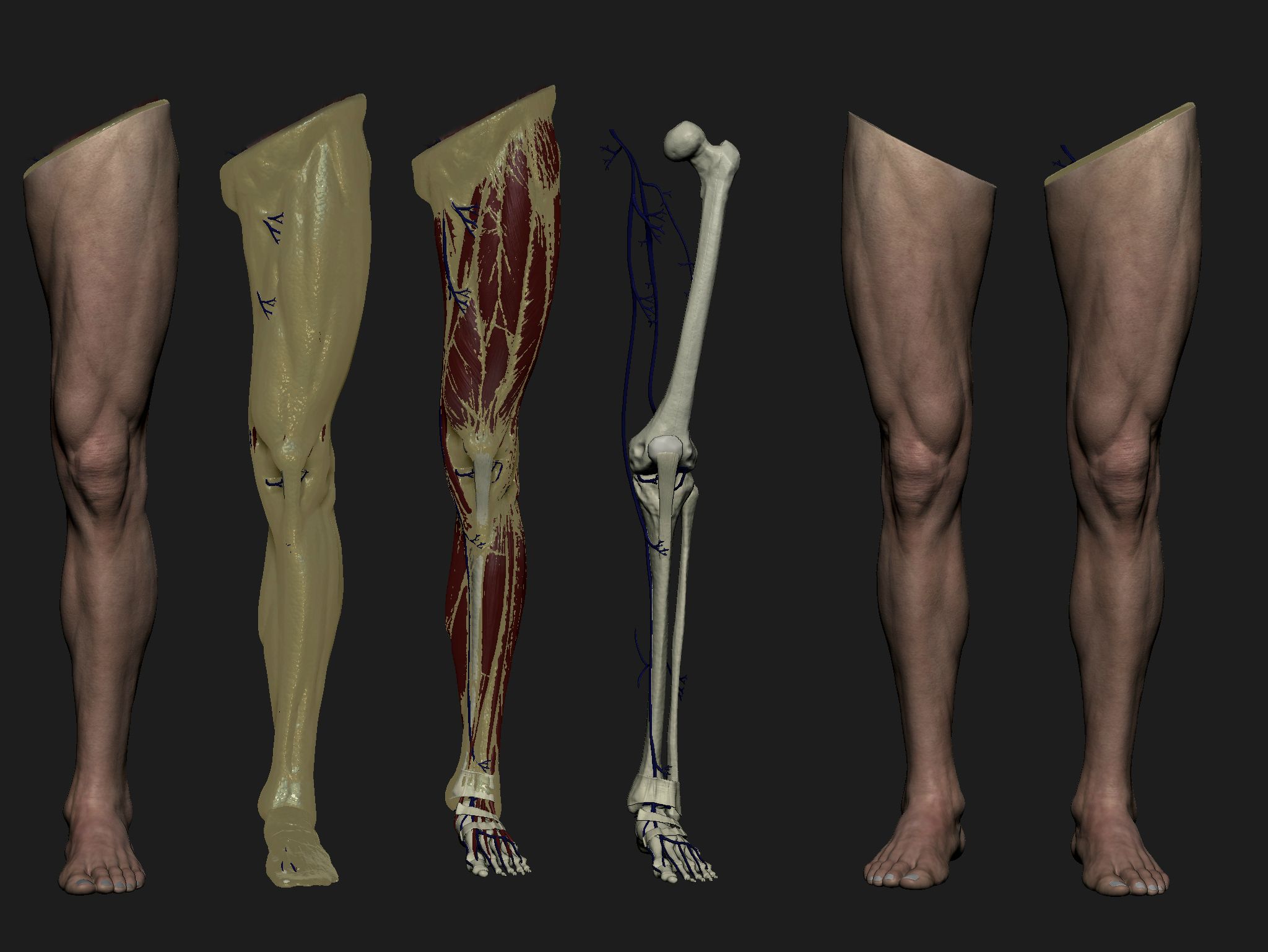 Она представлена двумя подкожными венами — большая подкожная вена (БПВ — v. saphena magna) и малая подкожная вена (МПВ — v. saphena parva).
Она представлена двумя подкожными венами — большая подкожная вена (БПВ — v. saphena magna) и малая подкожная вена (МПВ — v. saphena parva).
БПВ начинается от внутренней краевой вены стопы и на всем протяжении принимает много подкожных ветвей поверхностной сети голени и бедра . Впереди внутренней лодыжки она поднимается на голень и огибая сзади внутренний мыщелок бедра, поднимается до овального отверстия в паховой области. На этом уровне она впадает в бедренную вену. Большая подкожная вена считается самой длинной веной в организме, имеет 5-10 пар клапанов, её диаметр на всем протяжении составляет от 3 до 5 мм. В некоторых случаях большая подкожная вена бедра и голени может быть представлена двумя и даже тремя стволами.
МПВ начинается от наружной краевой вены стопы, собирающей кровь главным образом с подошвы. Обогнув наружную лодыжку сзади, она поднимается по середине задней поверхности голени до подколенной ямки. Начиная с середины голени, малая подкожная вена располагается между листками фасции голени в сопровождении медиального кожного нерва икры.
Пластическая анатомия человека, четвероногих животных и птиц
В учебнике описывается пластическая анатомия человека и животных. Автор книги художник с медицинским образованием, благодаря чему изложен только тот материал, который имеет практическую ценность для художников. Книга написана художником, имеющим высшее медицинское образование, благодаря чему изложен только тот материал, который имеет практическую ценность для художников; этим и особенно изложением методики изображения на анатомических основах книга выгодно отличается от других руководств того же профиля.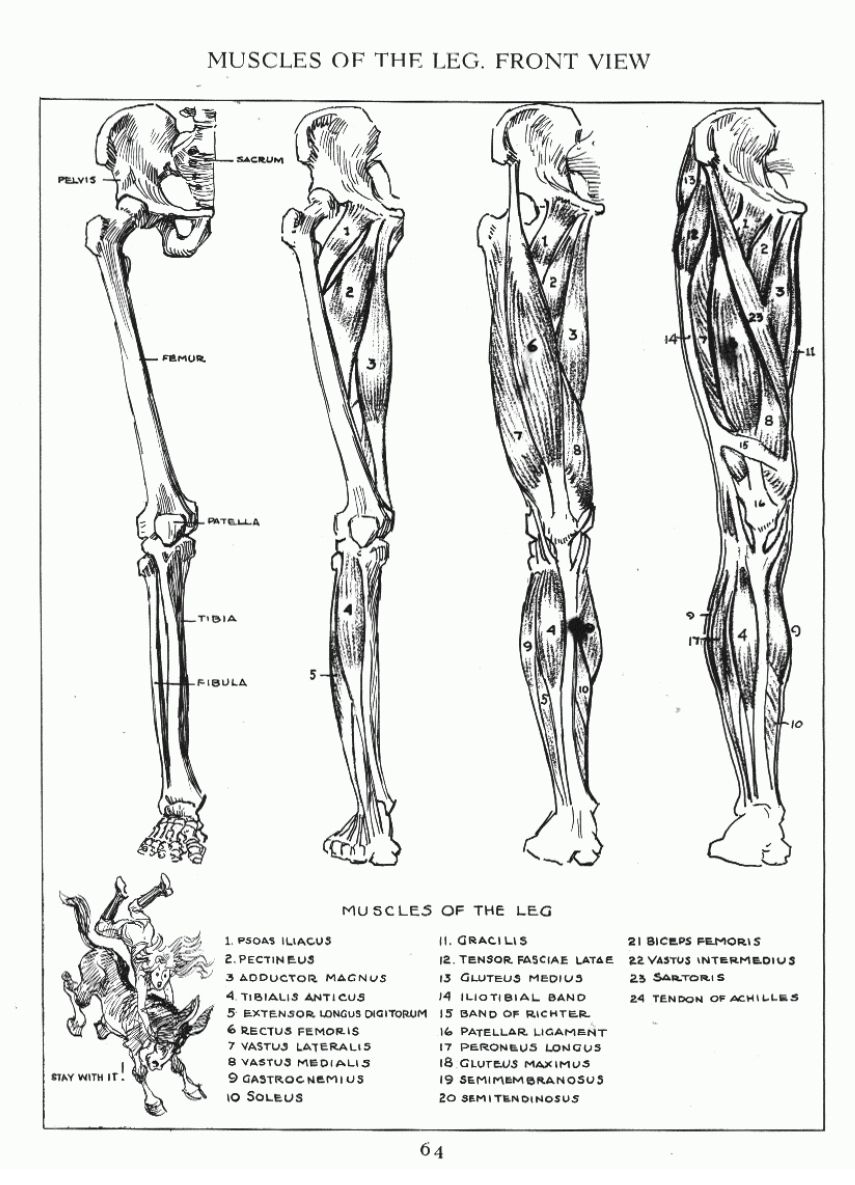 Книга содержит большое количество изображений: построение и схемы работы суставов, мышц и других органов, примеры построения фигуры, предварительные пластические обзоры участков тела и анатомический обзор восьми человеческих фигур в разных позах, а также разбираются классические рисунки лучших мастеров русской школы. Данный учебник хорошая база для изучения курса. Предназначается для студентов, изучающих изобразительное искусство. Может быть полезен в практической работе художника и послужить самоучителем для лиц, не изучавших пластическую анатомию.
Книга содержит большое количество изображений: построение и схемы работы суставов, мышц и других органов, примеры построения фигуры, предварительные пластические обзоры участков тела и анатомический обзор восьми человеческих фигур в разных позах, а также разбираются классические рисунки лучших мастеров русской школы. Данный учебник хорошая база для изучения курса. Предназначается для студентов, изучающих изобразительное искусство. Может быть полезен в практической работе художника и послужить самоучителем для лиц, не изучавших пластическую анатомию.
Укажите параметры рабочей программы
Дисциплина
Пластическая анатомия человека
УГС 43. 00.00 «СЕРВИС И ТУРИЗМ»
00.00 «СЕРВИС И ТУРИЗМ»
Специальность/профессия
Уровень подготовки
Суставы — анатомия и симптомы
Питание суставного хряща осуществляется с помощью внутри суставной жидкости, так в нем отсутствуют кровеносные сосуды
Существуют различные типы суставов:
- седловидные
- шаровидные и
- блоковидные
Это позволяет нам совершать разнообразные движения.
Существуют разные типы суставов: блоковидный (коленный, локтевой и между фалангами пальцев), цилиндрический (между лучевой и локтевой костью), седловидный (пястно-фаланговый сустав большого пальца кисти) и шаровидный (тазобедренный и плечевой).
Кости в суставах покрыты плотной хрящевой тканью. Волокна связок и сухожилий формируют вокруг сустава защитную оболочку — суставную капсулу.
Внутреннюю поверхность суставной капсулы выстилает синовиальная мембрана. Синовиальная мембрана выделяет в пространство сустава синовиальную жидкость, которая осуществляет питание хряща.
Суставной хрящ покрывает концы костей и устраняет небольшие неровности. Благодаря особому строению суставной хрящ поглощает ударные нагрузки.
Амплитуда движения зависит не только от типа сустава, но и от окружающих сустав структур (мышцы, связки, суставная капсула).
Артрит – воспаление сустава
Внутреннюю поверхность суставной капсулы выстилает синовиальная мембрана. Синовиальная мембрана выделяет в пространство сустава синовиальную жидкость, которая осуществляет питание хряща.
Синовиальная мембрана выделяет в пространство сустава синовиальную жидкость, которая осуществляет питание хряща.
Диагностика и лечение
Причины и симптомы проблем с суставами
Боль в суставах
Здоровый образ жизни
Узнайте о том, какой вид спорта подходит вам больше всего
Спорт и отдых
Анатомо-физиологический обзор стопы человека
Если у вас есть привилегия узнать кое-что, чего вы не знаете о своей стопе, воспользуетесь ли вы этой возможностью?
Если ваш ответ отрицательный, я предлагаю вам закрыть вкладку прямо сейчас, и если да, то давайте перейдем к обзору структуры и функции стопы.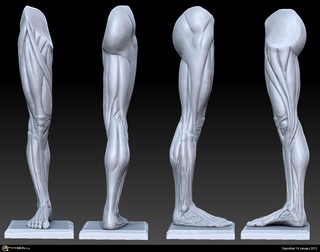
Стопа человека
Стопа человека состоит примерно из 26 костей, 33 суставов и более 100 связок, сухожилий и мышц.
Разве это не кажется излишним для этой плоской конструкции, прекрасно зная, что она выдерживает вес всего тела? Это должно сказать вам, из чего состоит ступня и какую работу она выполняет.
Основная причина, по которой вы все еще можете стоять сегодня, заключается в том, что ваши ноги находятся в хорошем состоянии. Стопа отвечает за поддержку и баланс веса тела.
Поэтому необходимо, чтобы стопа оставалась здоровой и в отличном состоянии, чтобы вы могли иметь правильную осанку, когда стоите в разных положениях и под разными углами.
А теперь поговорим немного о строении стопы!
Обзор костей стопы
• Фаланги, которые представляют собой кости пальцев ног
• Плюсневые кости, которые проходят через плоскую часть стопы
• Клинописные кости, ладьевидная кость и кубовидная кость, все из которых служат для создания твердой, но в некоторой степени гибкой основы.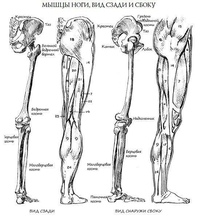
• Пяточная кость, которая является костью вашей пятки
• Таранная кость, которая является костью вашей лодыжки
• Таранная кость соединяется с большеберцовой костью, которая является основной костью голени.
Человеческая ступня делится на три категории: передняя, средняя и задняя.
Передняя часть стопы + средняя часть стопы
Передняя часть стопы состоит из пяти плюсневых костей и фаланг, известных как пальцы ног. Самая короткая из них — первая плюсневая кость, которая также является самой толстой и играет важную роль при движении вперед.
Ближе к головке первой плюсневой кости находятся две сесамовидные кости, которые соединены сухожилиями и связками.
Вторая, третья, четвертая и пятая плюсневые кости переднего отдела стопы более стабильны, потому что они защищены и не подвергаются чрезмерным тяговым усилиям.
Середина стопы представляет собой пирамидальную совокупность костей, которые составляют свод стопы.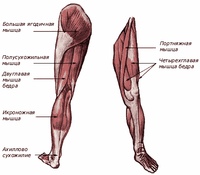 Он включает в себя пять из семи костей предплюсны, известных как ладьевидная, кубовидная и три клинопись.
Он включает в себя пять из семи костей предплюсны, известных как ладьевидная, кубовидная и три клинопись.
Передняя и средняя части стопы соединяются в пяти предплюсневых суставах.
Задняя стопа
Задняя часть стопы образует лодыжку и пятку и состоит из двух больших костей стопы, известных как таранная кость и пяточная кость.
Самая крупная кость предплюсны — это пяточная кость, она образует пятку, а таранная кость — стержень лодыжки.
Своды стопы
Две основные функции стопы — это выдерживать вес тела и двигаться. Для выполнения обоих сервисов он должен быть гибким и поддерживать высокую степень стабильности.
Стопа человека имеет три дуги, такие как:
Медиальная продольная дуга: она состоит из пяточной кости, ладьевидной кости, таранной кости, клинописи и трех плюсневых костей и очень важна, чем остальные две.
Боковая продольная дуга: состоит из пяточной кости, кубовидной кости, четвертой и пятой плюсневых костей.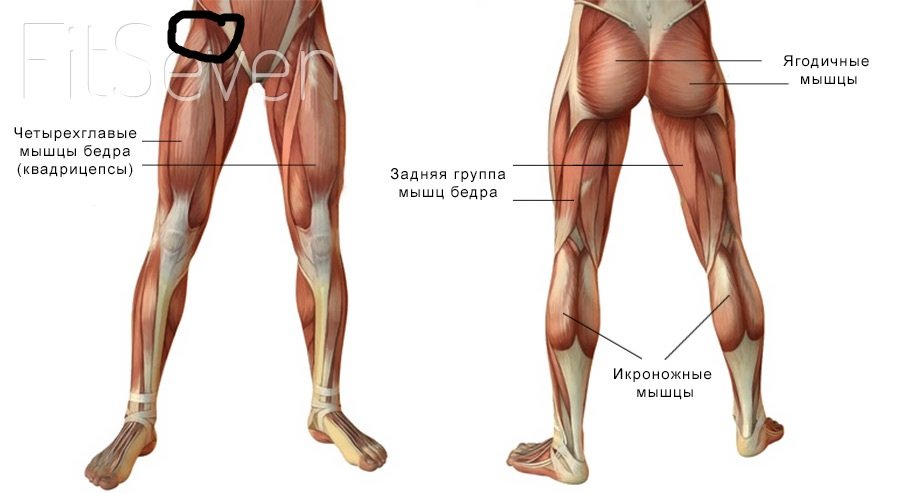
Поперечная дуга: состоит из клинописи, кубовидной формы, а также пяти плюсневых оснований стопы, известных как пальцы.
Связки стопы
Связка — это гибкая ткань, которая соединяет две кости вместе. Они прочные, и их довольно сложно повредить. Единственное повреждение связок — это растяжение связок, то есть разрыв или износ связок.
Но травмы также могут быть более опасными, и когда они случаются, это становится проблемой, потому что связки не получают кровоток, как кости и мышцы, и поэтому они очень медленно восстанавливаются.
Мышцы стопы
Стопа не только состоит из связок, но также имеет подчиненных, и важнейшей из них являются мышцы.
Конечно, в стопе много мышц, и их можно разделить на четыре категории, такие как внутренние мышцы подошвы, мышцы тыла (верхняя часть стопы), боковая мышца подошвы и медиальные мышцы стопы. единственным.
Сухожилие стопы
Подобно тому, как связки соединяют два сустава, существует также толстая полоса, которая соединяет мышцы с костями, и называется сухожилиями.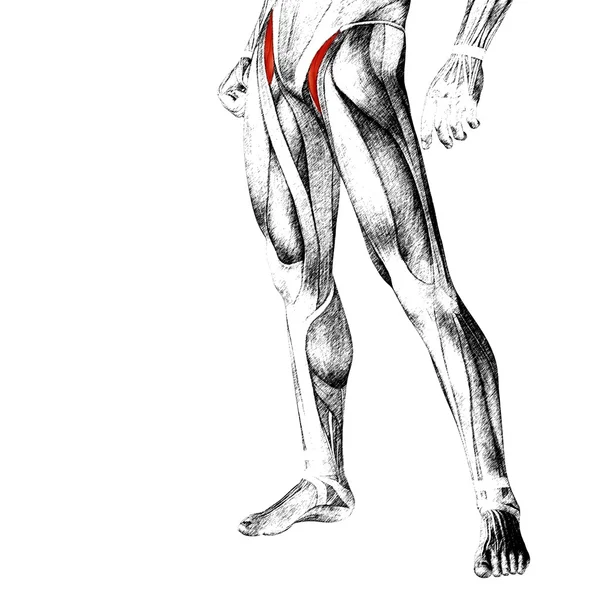 Он отвечает за движение, возникающее всякий раз, когда мышцы тянут кости.
Он отвечает за движение, возникающее всякий раз, когда мышцы тянут кости.
Состояние стопы
Подошвенный фасциит: возникает в результате воспаления связки подошвенной фасции вокруг нижней части стопы. Сопровождается болью в пятке и своде.
Стопа спортсмена: возникает в результате инфекции стопы, приводящей к сухой, шелушащейся, красной и раздраженной коже.
Травма ахиллова сухожилия: это связано с болью в задней части пятки и может указывать на проблему с ахилловым сухожилием.
Пяточная шпора: аномальный рост кости в пятке, который может вызывать сильную боль при ходьбе или стоянии. У людей с подошвенным фасциитом, плоскостопием или высоким сводом стопы более вероятно развитие пяточной шпоры.
Упавший свод стопы (плоскостопие): можно исправить ортопедической обувью.
Метатарсагал: это вызвано перегрузкой стопы и ношением неподходящей обуви, что сопровождается болью и воспалением в подушечке стопы.
Перелом: кость, которая может повредить кость, является плюсневой костью в результате травмы или многократного использования. Сопровождается припухлостью, болью, покраснением и синяками.
Сопровождается припухлостью, болью, покраснением и синяками.
Остеоартрит стопы: он возникает в результате износа компонентов стопы, а также в результате возраста. Сопровождается болью и отеками в стопах.
Средства для ухода за стопами
Ортопедические приспособления: это помогает исправить и лечить некоторые деформации стоп, поскольку они сконструированы таким образом, чтобы обеспечивать оптимальную поддержку стоп. Примерами являются туфли и сандалии с поддержкой свода и пятки.
Физическая терапия: она включает в себя множество физических нагрузок, требующих силы и поддержки ступней и лодыжек.
Хирургия стопы: в некоторых случаях правильным выбором является хирургическая операция.
Кости стопы — предплюсны — плюсны — фаланги
Кости стопы обеспечивают механическую поддержку мягких тканей; помогая ноге выдерживать вес тела в положении стоя и в движении.
Их можно разделить на три группы:
- Тарсалы — набор из семи костей неправильной формы.
 Они расположены проксимально в стопе в области щиколотки.
Они расположены проксимально в стопе в области щиколотки. - Плюсны — соединяют фаланги с предплюсневыми костями. Их пять — по одному на каждую цифру.
- Фаланги — кости пальцев ног. На каждом пальце ноги есть три фаланги — проксимальная, промежуточная и дистальная (кроме большого пальца ноги, у которого всего две фаланги).
Стопу можно также разделить на три области : (i) задняя часть стопы — таранная и пяточная кость; (ii) средняя часть стопы — ладьевидная, кубовидная и клинопись; и (iii) передняя часть стопы — плюсневые кости и фаланги.
В этой статье мы рассмотрим анатомию костей стопы — их костные ориентиры, суставы и клинические корреляции.
Рис. 1.0 — Обзор костей человеческой стопы. [/ caption]Тарсалс
Плюсневые кости стопы организованы в три ряда: проксимальный, промежуточный и дистальный.
Проксимальные кости предплюсны — таранная кость и пяточная кость.Они составляют заднюю часть стопы, образуя костный каркас вокруг проксимальных отделов лодыжки и пятки.
Талус
Таранная кость — самая верхняя из костей предплюсны. Он передает на стопу вес всего тела. Имеет три сочленения:
- Вверху — голеностопный сустав — между таранной костью и костями голени (большеберцовая и малоберцовая костей).
- Внизу — подтаранный сустав — между таранной костью и пяточной костью.
- Спереди — таранно-ладьевидный сустав — между таранной костью и ладьевидной костью.
Основная функция таранной кости — передавать силы от большеберцовой кости к пяточной кости (известной как пяточная кость). Он шире спереди по сравнению с сзади, что обеспечивает дополнительную стабильность лодыжки.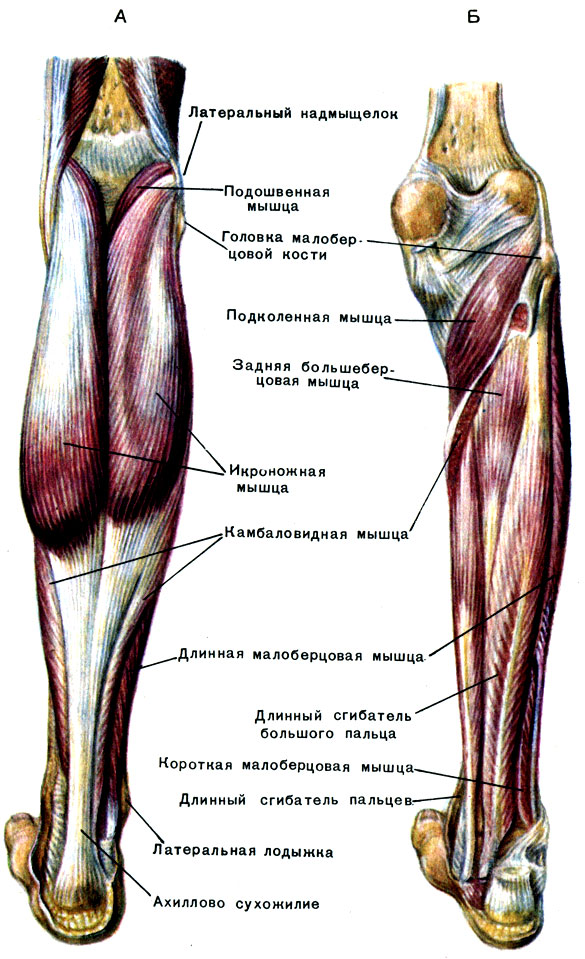
Несмотря на то, что к таранной кости прикрепляются многочисленные связки, никакие мышцы не берут начало и не прикрепляются к ней. Это означает, что существует высокий риск аваскулярного некроза , поскольку кровоснабжение сосудов зависит от фасциальных структур.
Calcaneus
пяточная кость является самой большой предплюсневой костью, а лежит под таранной костью, где она составляет пятку. Имеет два сочленения:
- Вверху — подтаранный (таранно-пяточный) сустав — между пяточной костью и таранной костью.
- Передняя часть — пяточно-кубовидный сустав — между пяточной костью и кубовидной.
Он выступает назад и принимает на себя вес тела, когда пятка ударяется о землю при ходьбе.Задняя часть пяточной кости обозначена бугристостью пяточной кости , к которой прикрепляется ахиллово сухожилие.
Рис.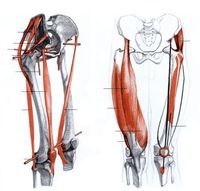 2. Кости предплюсны стопы. [/ caption] Промежуточная группа (Середина стопы)
2. Кости предплюсны стопы. [/ caption] Промежуточная группа (Середина стопы) Промежуточный ряд костей предплюсны содержит одну кость, ладьевидную кость (названную так потому, что она имеет форму лодки) .
Расположенный медиально, он сочленяется с таранной костью сзади, со всеми тремя клиновидными костями спереди и кубовидной костью сбоку.На подошвенной поверхности ладьевидной кости имеется бугорок для прикрепления части сухожилия задней большеберцовой мышцы .
Дистальная группа (средняя часть стопы)В дистальном ряду четыре предплюсневые кости — кубовидная и три клинописи. Эти кости сочленяются с плюсневыми костями стопы
. Кубовид является наиболее дальним латеральным, лежит впереди пяточной кости и позади четвертой и пятой плюсневых костей. Как следует из названия, он имеет форму куба.На нижней (подошвенной) поверхности кубовидной кости имеется бороздка для сухожилия длинной малоберцовой мышцы.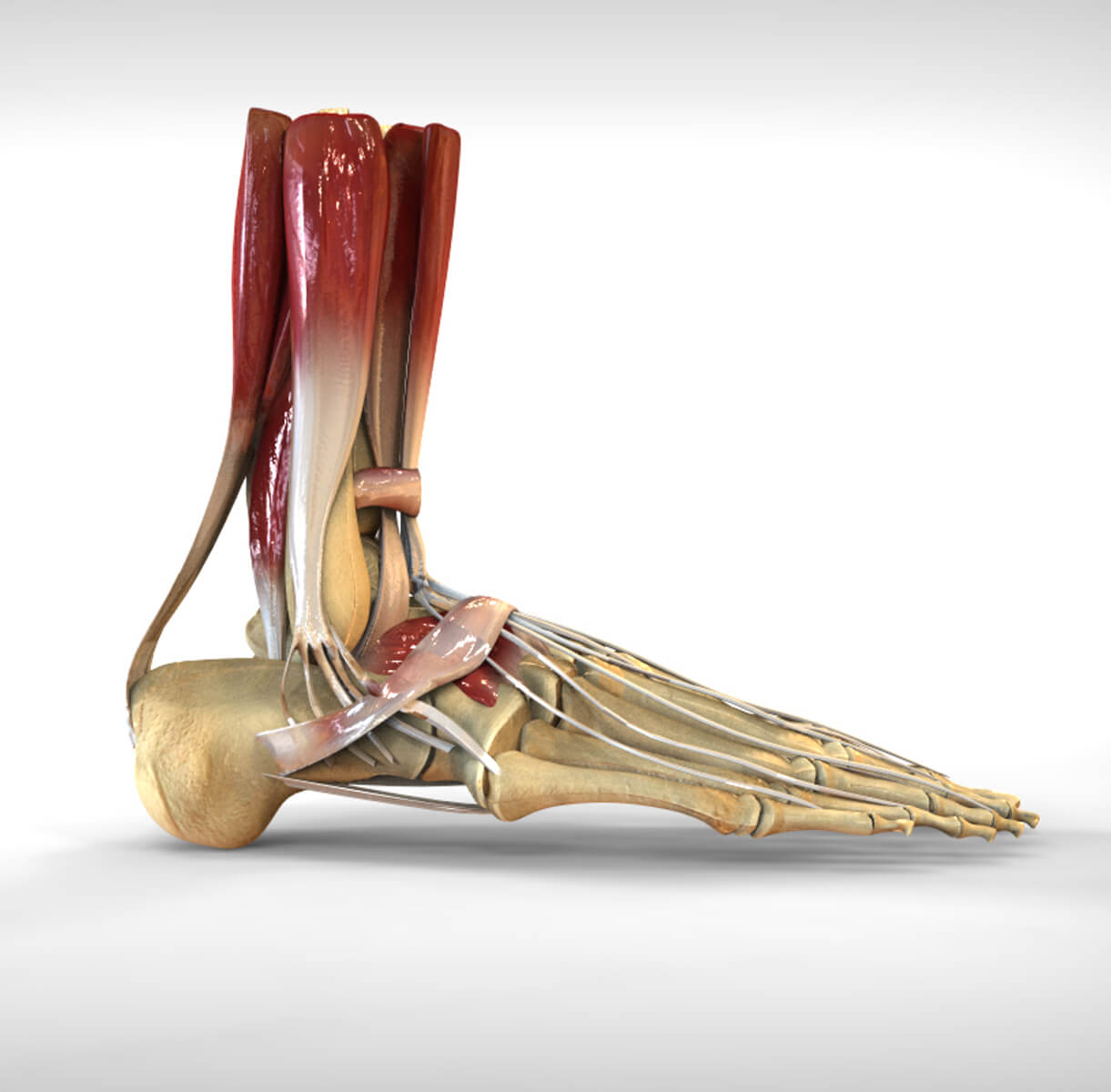
Три клинописи (боковые, промежуточные (или средние) и медиальные) представляют собой кости клиновидной формы. Они сочленяются ладьевидной костью сзади и плюсневыми костями спереди. Форма костей помогает сформировать поперечный свод стопы. Они также являются точкой крепления нескольких мышц:
- Медиальная клиновидная форма — передняя большеберцовая мышца, (часть) задняя большеберцовая мышца и длинная малоберцовая мышца
- Боковая клиновидная форма — короткий сгибатель большого пальца стопы
[начало клинической практики]
Клиническая значимость: переломы таранной и пяточной кости
Таранная кость и пяточная кость расположены в проксимальной части стопы и голеностопного сустава и участвуют в передаче сил от тела к земле.Это наиболее часто встречающиеся переломы костей предплюсны.
Талус
Переломы таранной кости чаще всего возникают в шейке таранной кости (50%), но могут возникать в теле таранной кости или латеральном отростке, причем переломы в головке таранной кости встречаются реже.
- Переломы шеи — обычно высокоэнергетические травмы, вызванные чрезмерным тыльным сгибанием стопы. Шейка таранной кости прижимается к большеберцовой кости. При этом типе перелома может нарушиться кровоснабжение таранной кости, что приводит к бессосудистому некрозу кости.
- Переломы тела — обычно возникают при прыжках с высоты.
Calcaneus
Пяточная кость часто ломается в результате осевой нагрузки , обычно в результате падения с высоты. Эти пациенты должны быть обследованы на предмет сопутствующих травм, таких как переломы поясничного отдела позвоночника или нижней конечности.
Кость может расколоться на несколько частей — оскольчатый перелом . На рентгеновском снимке пяточная кость будет казаться короче и шире.
Даже после лечения перелом пяточной кости может вызвать дополнительные проблемы. Подколенный сустав обычно разрушается, в результате чего сустав становится артритом . Пациент будет испытывать боль при перевороте и вывороте, что может сделать ходьбу по неровной поверхности особенно болезненной. Также может быть поражен пяточно-кубовидный сустав.
Пациент будет испытывать боль при перевороте и вывороте, что может сделать ходьбу по неровной поверхности особенно болезненной. Также может быть поражен пяточно-кубовидный сустав.
[окончание клинической]
плюсныПлюсневые кости расположены в передней части стопы , между предплюсневыми костями и фалангами.Они пронумерованы I-V (от медиального к латеральному).
Каждая плюсневая кость имеет похожее строение. Они выпуклые на дорсальной стороне и состоят из головы, шеи, стержня и основания (от дистального к проксимальному).
У них три или четыре сочленения:
- Проксимально — предплюсневые суставы — между основаниями плюсневых и предплюсневых костей.
- Латерально — межплюсневой сустав (и) — между плюсневой и прилегающей к ней плюсневой костью.
- Дистально — плюснефаланговый сустав — между головкой плюсневой кости и проксимальной фалангой.
[старт-клиника]
Клиническая значимость: переломы плюсневых костей
Переломы костей плюсны могут возникать по трем основным причинам.
Самый распространенный метод перелома — это прямой удар по стопе — обычно в результате падения тяжелого предмета на стопу.
Другой тип травмы плюсневой кости — это стрессовый перелом , неполный перелом, вызванный повторяющейся нагрузкой на кость. Это часто встречается у спортсменов и чаще всего возникает на шее второй и третьей плюсневой кости и проксимальной части пятой плюсневой кости.
Плюсневые кости также могут быть переломаны при чрезмерном выворачивании стопы. Если стопа сильно перевернута, короткая малоберцовая мышца может оторвать («оторвать») основание пятой плюсневой кости.
[окончание клинической]
Фаланги
фаланги — это кости пальцев ног. Все пальцы со второго по пятый имеют проксимальную, среднюю и дистальную фаланги. У большого пальца всего 2 пальца; проксимальная и дистальная фаланги.
У большого пальца всего 2 пальца; проксимальная и дистальная фаланги.
По строению они похожи на плюсневые кости, каждая фаланга состоит из основания, стержня и головы.
В чем уникальность стопы человека?
Наши ноги поддерживают нас. Кости, из которых состоят ступни, составляют четверть человеческого скелета, и тем не менее, несмотря на то, что они составляют такой большой процент тела, до недавнего времени они в значительной степени ускользнули от нас в летописи окаменелостей. Это расстраивает, потому что ясно, что эта история — эволюция человеческой стопы — захватывала нас на сотни лет. В конце концов, это связано с поворотным моментом в развитии двуногости в нашей истории.Понимание различий между нашими ногами и ногами других обезьян (как древних, так и современных) может дать нам ключ к разгадке изменений, которые были необходимы для двуногости — и, возможно, самого двуногого. То, что возникло, — это история разнообразия движений, которая поддерживает аргументы в пользу эволюции мозаики, что делает историю стопы в целом (не только нашей) замечательной.
В недавней обзорной статье исследователей Эллисона МакНатта и его коллег прослеживается литература по эволюции стопы человека.Поиски познания себя начинаются в 1699 году с анатомической оценки современных шимпанзе Эдвардом Тайсоном, который назвал их четырехчеловеческими, что означает, что все их придатки были приспособлены для работы в качестве рук. В 1863 году Томас Х. Хаксли сравнил ноги горилл и заметил, что, хотя они также были перевернутыми и обладали склонностью к хватанию, они также имели сходство мускулов с ногами человека. В 1935 году профессор анатомии Дадли Мортон предположил, что современная человеческая стопа является результатом двух различных переходов.В первом случае ступня должна была бы обладать более «обезьяноподобными» качествами с большей способностью к хватанию и гибкостью, и особенно с удлиненной областью средней части стопы. На более поздних стадиях стопа отошла бы от этих черт, хотя, возможно, сохранила способность «хватать» большим пальцем.
Идеи были хороши, но нам нужны были окаменелости. Открытие OH 8 в 1960 году командой Лики в Олдувайском ущелье продвинуло нас вперед. ОН 8 относится к Олдувайскому гоминиду номер 8 и датируется примерно 1.8 миллионов лет. Он принадлежал представителю семейства Homo habilis и включает левую предплюсневую и плюсневую кости — предплюсны — это серия костей, которые составляют плоскость ступни, ведущую к пальцам, но не пальцы. Вместе с открытием Люси, а также следами Лаэтоли, OH 8 послужил историей о том, что человеческая нога была адаптирована из ноги древесного шимпанзе. Однако в 1995 году ученые предположили, что анатомия голеностопного сустава и пятки (задней части стопы) существовала до анатомии передней части стопы человека, поскольку это относится к двуногому движению.Это было основано на открытии «Маленькой ноги», почти полного ископаемого скелета Australopithecus , извлеченного из Стеркфонтейна, Южная Африка, датируемого 3,3 миллиона лет назад, который демонстрировал аналогичные черты задней лапы. Ученые полагали, что ступня австралопитека была приспособлена для двуногого движения, но она также позволяла этому раннему предку человека при необходимости укрываться на деревьях.
Открытие OH 8 в 1960 году командой Лики в Олдувайском ущелье продвинуло нас вперед. ОН 8 относится к Олдувайскому гоминиду номер 8 и датируется примерно 1.8 миллионов лет. Он принадлежал представителю семейства Homo habilis и включает левую предплюсневую и плюсневую кости — предплюсны — это серия костей, которые составляют плоскость ступни, ведущую к пальцам, но не пальцы. Вместе с открытием Люси, а также следами Лаэтоли, OH 8 послужил историей о том, что человеческая нога была адаптирована из ноги древесного шимпанзе. Однако в 1995 году ученые предположили, что анатомия голеностопного сустава и пятки (задней части стопы) существовала до анатомии передней части стопы человека, поскольку это относится к двуногому движению.Это было основано на открытии «Маленькой ноги», почти полного ископаемого скелета Australopithecus , извлеченного из Стеркфонтейна, Южная Африка, датируемого 3,3 миллиона лет назад, который демонстрировал аналогичные черты задней лапы. Ученые полагали, что ступня австралопитека была приспособлена для двуногого движения, но она также позволяла этому раннему предку человека при необходимости укрываться на деревьях.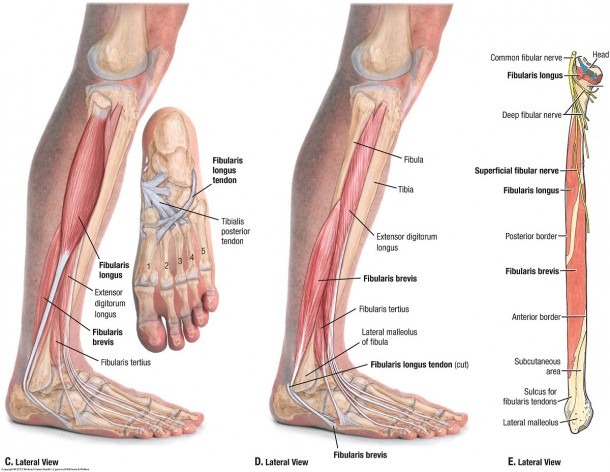 Объединив эти открытия по кусочкам, стало ясно, что эволюционная история человеческой стопы не может быть объяснена линейно.Человеческая нога развивалась независимо от других процессов эволюции человека и с разной скоростью у разных видов.
Объединив эти открытия по кусочкам, стало ясно, что эволюционная история человеческой стопы не может быть объяснена линейно.Человеческая нога развивалась независимо от других процессов эволюции человека и с разной скоростью у разных видов.
Одно из фактов, которые мы можем сказать с уверенностью, это то, что современная человеческая ступня не произошла от ступни шимпанзе. Как бы разумно ни казалось проводить сравнения между ними, явное расхождение между родами от последнего общего предка (LCA) означает, что гоминины и панины развили ноги в соответствии со своими потребностями. Основное различие между ними заключается в гибкости.Стопа первого приспособлена для жесткого отталкивания, необходимого для двуногого передвижения. Стопы последнего обладают большей гибкостью в целом и способностью к хватанию, что позволяет лазать по деревьям, а также двигаться на четвероногих ногах по земле. Тщательный анатомический анализ показывает, что многие из различий между ними связаны с жесткостью одного по сравнению с гибкостью другого с сильными и изящными чертами, которые поддерживают разную мускулатуру и движения.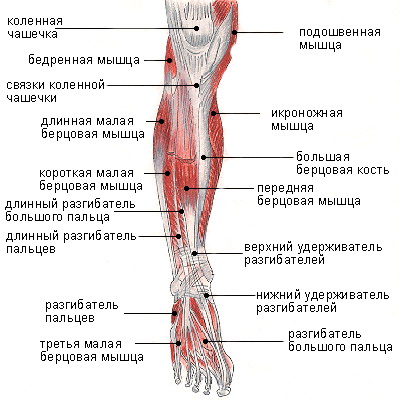 Например, у людей большой палец ноги толще, чем у шимпанзе, и он совмещен с остальными пальцами, что позволяет ноге отталкиваться от земли.Этот «большой палец ноги» не только более изящен у шимпанзе, но и изгибается по направлению к другим пальцам, обеспечивая более сильное сгибание. Эти характеристики относятся и к пальцам в целом. У людей эти кости более прочные и могут помочь поглотить часть давления отталкивания, в то время как у шимпанзе эти кости вытянуты и изогнуты с большей степенью гибкости.
Например, у людей большой палец ноги толще, чем у шимпанзе, и он совмещен с остальными пальцами, что позволяет ноге отталкиваться от земли.Этот «большой палец ноги» не только более изящен у шимпанзе, но и изгибается по направлению к другим пальцам, обеспечивая более сильное сгибание. Эти характеристики относятся и к пальцам в целом. У людей эти кости более прочные и могут помочь поглотить часть давления отталкивания, в то время как у шимпанзе эти кости вытянуты и изогнуты с большей степенью гибкости.
Нога шимпанзе, скорее всего, содержит производные элементы, которые были адаптированы для поддержания их древесного образа жизни.Если мы хотим открыть «стопу предков», современная человеческая ступня может содержать подсказки. Мы еще не определили последнего общего предка, которого мы разделяли с шимпанзе, но у нас есть очень старые окаменелости гоминидов, такие как Pierolapithecus catalaunicus (11,9 миллиона лет) и ископаемые останки гомининов, такие как Ardipithecus ramidus (4,4 миллиона лет). Последняя — самая старая из имеющихся у нас окаменелостей гомининов с относительно полной стопой. Между этими подставками для книг и другими обезьянами миоцена мы можем сделать то же самое, что и Мортон, и создать предложение для LCA.Мы можем предсказать, что он мог обладать грациозными чертами, облегчающими хватание и гибкостью. Он мог бы быть перевернут, но он также мог пройти через более жесткую и прочную среднюю часть стопы, чтобы обеспечить возможность земной активности.
Последняя — самая старая из имеющихся у нас окаменелостей гомининов с относительно полной стопой. Между этими подставками для книг и другими обезьянами миоцена мы можем сделать то же самое, что и Мортон, и создать предложение для LCA.Мы можем предсказать, что он мог обладать грациозными чертами, облегчающими хватание и гибкостью. Он мог бы быть перевернут, но он также мог пройти через более жесткую и прочную среднюю часть стопы, чтобы обеспечить возможность земной активности.
После перехода от Australopithecus к Homo пальцы ног уменьшились в длине и кривизне, лодыжка и соответствующая мускулатура уменьшились в размерах, и появились полные своды стопы. Большой палец ноги сместился, чтобы выровняться с другими пальцами, а не загибаться внутрь, что позволило более эффективно отталкиваться для двуногости.Есть некоторые исключения из этих разработок. У Homo naledi , например, пальцы ног более изогнуты, чем у всего рода Homo. А Homo floriensis имеет удлиненную переднюю часть стопы, которая больше всего напоминает лапку бонобо. В таких вариациях нет ничего необычного; пяточные кости современных человекообразных обезьян различаются в зависимости от вида. Эти случаи иллюстрируют разнообразие эволюции стопы и передвижения, которые со временем могут дать более контекстуальные подсказки о жизни этих групп.
В таких вариациях нет ничего необычного; пяточные кости современных человекообразных обезьян различаются в зависимости от вида. Эти случаи иллюстрируют разнообразие эволюции стопы и передвижения, которые со временем могут дать более контекстуальные подсказки о жизни этих групп.
История человеческой стопы все еще разворачивается. Он уникален тем, что лучше всего соответствует нашему стилю двуногого передвижения. Изменения, обнаруженные учеными в костях стопы австралопитов, предполагают, что существовала разница в том, как они ходили даже между собой, что верно для людей сегодня: у нас разная длина шага и разные способы опускаться на ноги. У некоторых более сильный шаг, чем у других, не говоря уже о том, как ступни танцоров меняются с годами тренировок.В нашей истории есть дополнительная сложность, связанная с воздействием привычного ношения обуви. Это изменило то, как мы ходим, и, несомненно, изменило морфологию наших ног. При этом необходимо будет учитывать возможности протезирования и доступности. История, хотя и неполная, остается не менее увлекательной, чем это было сотни лет назад, когда у нас была только сравнительная оценка.
История, хотя и неполная, остается не менее увлекательной, чем это было сотни лет назад, когда у нас была только сравнительная оценка.
Комментарии здесь отключены. Но вы можете присоединиться к сообществу на Facebook.
–
Ссылка:
McNutt EJ, Zipfel B, DeSilva JM.Эволюция человеческой стопы. Evol Anthropol. 2018; 1–21. https://doi.org/10.1002/evan.21713
–
Возможно вам понравится:
У древней девочки были денисовцы и неандертальцы.
Наши отношения с хлебом, которым уже 14 400 лет.
Почему люди хотят пить воду из саркофага?
Мы изменили наше поведение, чтобы мы могли писать и ходить.
Обертывания показывают, что мумификация в Египте началась раньше, чем мы думали.
Что зубы могут рассказать нам о наших доисторических предках?
Разгадывая правду в записях о продуктах питания
Переосмысливая эволюцию стопы человека: выводы из экспериментальных исследований
РЕЗЮМЕ
Адаптивные объяснения анатомии современной человеческой стопы долгое время интересовали биологов-эволюционистов из-за резких различий между нашими ногами и ногами. ближайшие живущие родственники, человекообразные обезьяны.Морфологические особенности, включая галлюкальную противоположность, длину пальцев и продольный свод стопы, традиционно использовались для дихотомии ступней человека и большой обезьяны как приспособленных для двуногой ходьбы и передвижения по деревьям соответственно. Однако недавние биомеханические модели функции стопы человека и экспериментальные исследования передвижения больших обезьян подорвали эту простую дихотомию. Здесь мы рассматриваем это исследование, уделяя особое внимание биомеханике удара стопой, отталкивания и накопления упругой энергии в стопе, и показываем, что люди и человекообразные обезьяны имеют некоторые недооцененные, удивительные сходства в функциях стопы, например, в использовании стопы и способности укрепить среднюю часть стопы.Мы также показываем, что некоторые уникальные особенности человеческой стопы, включая пружинный продольный свод и короткие пальцы ног, вероятно, являются адаптацией к бегу на длинные дистанции. Мы используем эту схему для интерпретации летописи окаменелостей и утверждаем, что человеческая ступня прошла через три стадии эволюции: во-первых, ступня огромной обезьяны, приспособленная для передвижения по деревьям, но с некоторыми приспособлениями для двуногой ходьбы; во-вторых, ступня, приспособленная для эффективной двуногой ходьбы, но сохранившая некоторые древесные хватательные приспособления; и в-третьих, человекоподобная ступня, приспособленная для повышенной экономичности при ходьбе и беге на длинные дистанции, утратившая цепкость.Основываясь на этом сценарии, мы предполагаем, что отбор на двуногий бег сыграл важную роль в потере древесных приспособлений.
Введение
Человеческие ноги отличаются от ног других животных, включая наших ближайших ныне живущих родственников, человекообразных обезьян [в этой статье мы используем термин «человекообразные обезьяны» для обозначения парафилетической группы, включающей всех существующих членов семейства Hominidae кроме людей (например, шимпанзе, бонобо, гориллы и орангутанги)], во многих характеристиках, связанных с нашей уникальной формой двуногого передвижения.К ним относятся большая пяточная кость, короткие пальцы ног, приведенная и не противопоставленная большая палец (см. Глоссарий), а также хорошо развитые продольные и поперечные дуги (см. Глоссарий; Рис. 1A). Большинство адаптивных объяснений этих особенностей основано на наблюдаемых различиях в функциях ног человека и обезьяны во время передвижения. Люди используют свои ступни в качестве жестких рычагов во время двуногой ходьбы, в то время как ступни больших обезьян, кажется, сгибаются в области средней части стопы (см. Глоссарий) во время отталкивания (см. Глоссарий), предположительно из-за отсутствия у них сводов стопы и подвижных суставов средней части стопы для повышения схватывания во время движения. древесное передвижение (см. Глоссарий; рис.1B) (DeSilva, 2010; Elftman, Manter, 1935a). Другие исследования документально подтвердили, как человекообразные обезьяны используют свои длинные пальцы ног, противоположные галлюцинации и подвижные лодыжки для захвата древесных опор (DeSilva, 2009; Holowka et al., 2017a; Morton, 1924). Эти наблюдения лежат в основе того, что стало консенсусной моделью эволюции стопы человека: выбор для двуногой ходьбы произошел за счет древесных локомоторных способностей, что привело к дихотомии между анатомией и функцией стопы человека и большой обезьяны. Согласно этому образу мышления предполагается, что анатомические особенности стопы, характерные для человекообразных обезьян, представляют собой приспособления к древесному поведению, а те, которые уникальны для человека, считаются связанными с двуногой ходьбой.
Этой дихотомией долгое время руководствовались реконструкции передвижения ископаемых гомининов (см. Глоссарий) и, в более широком смысле, теории эволюции двуногого человека. Все ступни ранних гомининов демонстрируют различную мозаику человеческих и обезьяноподобных черт; следовательно, локомоторное поведение ранних гомининов обычно реконструируется как переходное между людьми и человекообразными обезьянами. Из-за предполагаемого компромисса между древесным и двуногим передвижением, виды с большими обезьяноподобными ногами считаются менее эффективными двуногими, а виды с более похожими на человека ногами считаются менее древесными.Эта структура часто приводит к противоречивым реконструкциям передвижения ископаемых гомининов. Например, кости стопы раннего гоминина Australopithecus afarensis были интерпретированы как указывающие на то, что этот вид либо сохранил приспособления к лазанию, что поставило под угрозу способность двуногой ходьбы (Harcourt-Smith and Aiello, 2004; Susman et al., 1984), либо был современным человекоподобные двуногие больше не зависят от древесных движений (Latimer, Lovejoy, 1989; Ward et al., 2011). Это противоречие представляет собой слишком распространенный тупик в реконструкциях передвижения ископаемых гомининов, в которых доминирующая интерпретирующая структура основана на дихотомии человек-большая обезьяна, препятствуя интерпретации эволюции сложных и уникальных человеческих черт, связанных с двуногостью.
Одной из основных проблем этой дихотомии является предположение о том, что все особенности человеческой стопы являются адаптациями для двуногой ходьбы, что не учитывает возможность того, что человеческая ступня может быть адаптирована для поведения, не связанного с ходьбой, такого как бег (Bramble and Lieberman, 2004 ). Кроме того, предполагается, что лапы большой обезьяны приспособлены в первую очередь для передвижения по деревьям, хотя наземное четвероногое движение является важным компонентом локомоторного поведения у всех африканских человекообразных обезьян (Doran, 1996), и нелокомоторное поведение, такое как борьба, также могло быть выбрано для (Кэрриер и Каннингем, 2017 г.).Вторая проблема заключается в аргументе замысла, что анатомические структуры у существующих видов с особым поведением могут быть использованы для вывода такого же поведения в окаменелостях, даже несмотря на то, что многие предполагаемые взаимосвязи между структурой и функцией имеют мало эмпирической поддержки при тестировании (Lauder, 1995). Решение этих проблем требует разработки и тестирования биомеханических моделей конкретных аспектов функции стопы. Если модель может быть эмпирически определена для поведения in vivo в соответствии с прогнозом, ее можно будет применить в более широком смысле для вывода функции из анатомии у живых и ископаемых видов.Этот подход, основанный на моделях, особенно важен для проверки гипотез о стопе, учитывая ее фундаментальную роль в передвижении и различия между ступнями человека, человекообразных обезьян и других млекопитающих. Базовая механика перемещения центра масс во время ходьбы и бега одинакова у млекопитающих с разным размером тела (Cavagna et al., 1977), но в то время как большинство млекопитающих контактируют с землей только пальцами (дигитгради) или кончиками пальцев ног. (унгулиграда), люди и человекообразные обезьяны стопоходят, опуская всю ступню вниз, включая пятку (рис.1) (Hildebrand, Goslow, 2001; Schmitt, Larson, 1995). Как показали недавние экспериментальные исследования, плантиграда (см. Глоссарий) в целом и позы стопы человека в частности имеют важные биомеханические последствия как при ударе, так и при отталкивании стопы во время ходьбы и бега.
Здесь мы используем основанную на модели интерпретирующую структуру для обзора эволюции стопы человека, сосредоточив внимание на трех основных кинематических и кинетических проблемах ходьбы и бега: (1) как стопа справляется с силами удара (см. Глоссарий), когда нижняя конечность изначально сталкивается с землей, (2) как ступня создает движущий рычаг для отталкивания и (3) как стопа накапливает и высвобождает упругую энергию во время бега.Мы используем этот Обзор для интерпретации новых данных экспериментальных исследований биомеханики стопы больших обезьян, а затем применяем эти идеи к летописи окаменелостей гомининов для оценки сил отбора, которые сформировали анатомию стопы человека и, в более широком смысле, эволюцию двуногого мышления. Поскольку мы фокусируемся на стопе, мы не покрываем непосредственно голеностопный сустав (голеностопный и подтаранный суставы), но мы отсылаем читателей к Pontzer et al. (2014), O’Neill et al. (2015) и Зелик и Адамчик (2016) за обзоры сравнительной биомеханики голеностопного сустава у людей и человекообразных обезьян.Кроме того, мы ограничиваем наш обзор исследованиями наземного передвижения, но рекомендуем Картмилла (1985), Ричмонда (2007), ДеСильва (2009), Венкатарамана и др. (2013), Holowka et al. (2017a) и Wunderlich и Ischinger (2017) за обсуждение биомеханики стопы приматов и схватывания за педали во время древесного передвижения. Наконец, мы признаем роль стопы в поведении, отличном от стационарного передвижения, которое может иметь серьезные последствия для физической формы, например в борьбе, маневрировании и ускорении.Этому поведению уделяется меньше внимания в исследованиях биомеханики стопы, но, тем не менее, их важно учитывать при понимании эволюции стопы, и поэтому мы отсылаем читателей к обсуждениям в Reeser et al. (1983), Carrier et al. (1994) и Кэрриер и Каннингем (2017).
ГлоссарийПередвижение по деревьям
Двигательное поведение, такое как лазание, которое связано с путешествием по веткам, стволам деревьев и другим растительным структурам.
Australopithecines
Ископаемые виды гомининов (см. Ниже), принадлежащие к роду Australopithecus , который существовал примерно 2–4 миллиона лет назад в Африке и предшествовал роду Homo .
Cercopithecines
Таксономическая группа, включающая большинство азиатских и африканских видов обезьян, включая макак и бабуинов. Они более тесно связаны с людьми и человекообразными обезьянами, чем южноамериканские обезьяны.
Потеря энергии при столкновении
Поскольку поддержка веса передается с одной ноги на другую во время двуногой ходьбы, скорость центра масс тела направляется вперед и вниз, а затем должна быть перенаправлена вперед и вверх.Потеря энергии при столкновении описывает механическую работу, которая должна быть выполнена для изменения скорости центра масс тела во время этого поэтапного перехода.
Бег на выносливость
Бег у людей, поддерживаемый в течение длительного периода времени в умеренном темпе при аэробном метаболизме.
Hallux
Первый луч педали, включая плюсневую кость и фаланги. У людей это то, что называют «большим пальцем ноги», а у нечеловеческих приматов большой палец стопы противопоставлен другим пальцам на педали, как большой палец с пальцами.
Гоминины
Люди и все ископаемые виды, которые более тесно связаны с людьми, чем шимпанзе.
Сила удара
Всплеск силы реакции опоры после первоначального контакта стопы, вызванный сохранением количества движения при быстром замедлении конечности.
Внутренние мышцы стопы
Мышцы, которые образуются и прикрепляются к костям стопы, включая отводящий большой палец, короткий сгибатель пальцев и квадратную мышцу подошвы.
Последний общий предок
Термин, используемый для описания последнего общего предка шимпанзе и человека, вида, от которого произошли линии, ведущие к шимпанзе и человеку, существовавшие примерно 6–9 миллионов лет назад. Обычно сокращенно LCA.
Продольный свод стопы
Уникальный для человека приподнятый участок стопы, проходящий в продольном направлении от пятки до пальцев ног. Он определяется костной структурой костей предплюсны и плюсны и поддерживается связками и мышцами.На медиальной стороне стопы он выше, чем на боковой.
Плюсно-фаланговые суставы
Суставы между головками плюсневых костей и проксимальными фалангами (костями пальцев).
Средняя часть стопы
Область стопы дистальнее пятки и голеностопного сустава, включая все суставы проксимальнее плюсне-фаланговых суставов. У людей продольный свод охватывает всю среднюю часть стопы.
Midtarsal break
Явление, наблюдаемое во время ходьбы по земле у человекообразных обезьян сразу после подъема пятки.Когда пятка отрывается от земли, стопа изгибается в области средней части стопы, так что часть средней части стопы на короткое время сохраняет контакт с землей.
Подошвенный апоневроз
Широкий лист связочной соединительной ткани с характерными латеральными и центральными частями, который прикрепляется проксимально к подошвенным пяточным клубням и дистально к основанию проксимальных фаланг и структурам мягких тканей под головками плюсневых костей.
Plantigrady
Поза стопы, используемая во время движения, когда вся подошвенная поверхность стопы, включая пятку, соприкасается с землей.
Отталкивание
Период второй половины фазы опоры шага ходьбы или бега у людей, начинающийся, когда пятка начинает подниматься от земли, и заканчивается, когда пальцы ног отрываются от земли.
Стойка
Фаза походки, когда ступня стоит на земле.
Поперечная дуга
Поднятая область стопы в поперечной плоскости сегмента, определяемая проксимально конформацией кубовидной и клиновидной костей, а дистальнее — плюсневыми костей.Он присутствует у людей и, как утверждается, присутствует, но в меньшем количестве, у человекообразных обезьян, хотя это различие не было определено количественно.
Механизм брашпиля
У людей тыльное сгибание пальцев стопы создает момент вокруг плюснефаланговых суставов, который преобразуется в линейную силу на подошвенный апоневроз. Это увеличивает напряжение подошвенного апоневроза и подтягивает пяточную кость к головкам плюсневых костей, поднимая продольный свод и укрепляя стопу.
Удар ступней и столкновение
Одной из характерных черт двуногой ходьбы человека является удар ступней, при котором пятка касается земли раньше остальной ступни.Человекообразные обезьяны также часто ударяются пяткой во время двуногой и четвероногой ходьбы (Elftman and Manter, 1935a; Schmitt and Larson, 1995), хотя иногда они касаются пяткой и боковой стороной стопы одновременно (Vereecke et al., 2003). Некоторые производные аспекты морфологии пяточной кости человека предполагают адаптацию к двуногому удару пяткой, включая крепкий пяточный бугорок и латеральный подошвенный отросток (Fig. 1A). Поскольку пяточная обезьяна без латерального подошвенного отростка считается слишком гибкой, чтобы выдерживать нагрузки, возникающие при двуногом движении (Latimer and Lovejoy, 1989), морфология пяточной кости была использована для реконструкции локомоции ископаемых гомининов (например.г. DeSilva et al., 2013). Однако в сравнительных исследованиях не было четко сформулировано и не проверено, препятствует ли изящная пяточная кость двуногому поведению, а также не выяснено, почему удар пяткой полезен при ходьбе, подобной человеческой.
Рис. 1.Ноги человека и шимпанзе. (A) Скелеты ступней человека и шимпанзе, вид сверху (вверху) и вид медиально (внизу). Острие стрелки указывает на верхнюю часть головы плюсневой кости, присутствующую у людей, но не у шимпанзе. (B) Общая кинематика ног человека и шимпанзе во время отталкивания при ходьбе на двух ногах.В середине у обоих видов подошвенная поверхность стопы ровная на земле. Эта поза называется «плантиградией». Впоследствии продольный свод стопы помогает превратить человеческую ступню в жесткий рычаг, позволяя одновременно поднимать пятку и среднюю часть стопы от земли во время отталкивания. У шимпанзе средняя часть стопы кратковременно сохраняет контакт с землей после подъема пятки из-за тыльного сгибания подвижных суставов средней части стопы. Это движение называется «разрывом средней предплюсны» и обозначается стрелкой.Изменено из книги Эльфтмана и Мантера (1935a).
Хорошо известно, что ходьба и бег на двух ногах с ударом пяткой влечет за собой временное быстрое увеличение вертикальной силы реакции опоры в начале стойки (см. Глоссарий), называемую ударной силой (рис. 2A, C). Эти ударные силы участвуют в повреждении тканей, приводящем к дегенеративным заболеваниям суставов и другим скелетно-мышечным травмам (Whittle, 1999), хотя биомеханические факторы, определяющие эти силы и то, как они передаются через тело, полностью не изучены.Недавно в нескольких исследованиях была разработана и протестирована биомеханическая модель удара стопой у людей, которая демонстрирует, что силы удара возникают, когда некоторая часть нижней конечности, эффективная масса, резко замедляется, что требует быстрого рассеивания механической энергии (Chi and Schmitt, 2005; Либерман и др., 2010). Согласно этой модели, пик удара является функцией скорости стопы при ударе, податливости голеностопного и коленного суставов и продолжительности замедления, которая сама зависит от сжатия толстой волокнистой жировой подушки ниже пяточного бугра (Gefen et al., 2001). Эта пяточная накладка рассеивает 17–45% энергии, возвращаемой от земли при ударе человека (Gefen et al., 2001; Pain and Challis, 2001). Человекообразные обезьяны часто создают ударную силу при двуногом ходьбе (рис. 2B) (Crompton et al., 2008; Pontzer et al., 2014), но также обладают толстыми подушечками на пятке, которые предположительно рассеивают некоторую энергию при ударе стопой (Raven, 1936; Vereecke и др., 2003). Кроме того, на основе описанной выше модели человекообразные обезьяны должны быть способны смягчать ударные силы во время двуногой ходьбы за счет уменьшения скорости стопы перед приземлением и приземлением с более гибкими положениями суставов.В частности, используя большие углы сгибания колена, человекообразные обезьяны потенциально могут снизить эффективную массу конечности, которая замедляется при ударе ступней.
Рис. 2.Вертикальные силы реакции опоры во время фазы опоры при двуногом передвижении. Красный указывает силу удара. Сила выражается в единицах массы тела (BW). И люди (A), и шимпанзе (B) производят пик удара после удара ногой во время двуногой ходьбы. Люди создают высокую пиковую силу удара при беге с ударом пятки (C), но обычно не создают пиковой силы удара при беге с ударом передней части стопы (D).Изображения стопы модифицированы по материалам Эльфтмана и Мантера (1935a).
Недавно Уэббер и Райхлен (2016) продемонстрировали, что люди могут уменьшить или избежать силы удара во время ходьбы, сначала коснувшись земли передней частью стопы. Кроме того, используя позы для пальцев во время ходьбы, люди могут увеличить длину нижней конечности от бедра до точки контакта с землей, что теоретически увеличит расстояние, пройденное за один шаг. Почему же тогда люди ходят с ударами пяток и стопой? Одна потенциальная причина заключается в столкновительной модели опоры центра масс: при ударе пяткой центр масс тела перемещается вперед и вниз, но должен быть перенаправлен вперед и вверх.Количество отрицательной работы, которую должна выполнить ведущая конечность, чтобы вызвать это изменение, называемое «потери энергии при столкновении» (см. Глоссарий), пропорционально квадрату угла между конечностями при ударе ступней (Donelan et al., 2002). Приземляясь на заднюю часть стопы (пятку) ведущей конечностью и отталкиваясь от передней части ступни (пальцев ног) задней конечностью, люди резко сокращают этот угол и, следовательно, стоимость ходьбы (Адамчик и Куо, 2013). Действительно, Cunningham et al.(2010) обнаружили, что люди должны выполнять больше работы, чтобы преодолеть потери при столкновении при ходьбе пальподобной, а не стопоходящей позе. Кроме того, они обнаружили, что плантиграда снижает внешние суставные моменты, которым должны сопротивляться голеностопные мышцы, по сравнению с дигитградией. Эта стратегия отличает людей от других наземных двуногих (Usherwood et al., 2012) и может частично объяснять относительно низкую метаболическую стоимость ходьбы у людей (Biewener et al., 2004; Cunningham et al., 2010).Кроме того, Уэббер и Райхлен (2016) продемонстрировали, что использование удара пяткой в начале стойки во время стопоходящей ходьбы позволяет центру давления перемещаться вперед под стопой во время фазы стойки, эффективно увеличивая длину шага и улучшая экономичность ходьбы человека.
Поскольку человекообразные обезьяны используют позу стопы, мы предполагаем, что они также извлекают выгоду из описанных выше преимуществ при двуногом движении. Однако, поскольку человекообразные обезьяны используют различные позы для ударов ног (Schmitt and Larson, 1995; Vereecke et al., 2003), мы могли бы предсказать, что у них самая длинная эффективная длина шага и самые низкие затраты на столкновение при приземлении с ударом пятки. Еще предстоит проверить, создают ли удары пяткой обезьяны более высокие силы удара, чем другие позы для ударов ног, как у людей (Webber and Raichlen, 2016).
Хотя удары пяткой энергетически выгодны при двуногой ходьбе, во время бега человека они вызывают силы удара и нагрузки примерно в два-четыре раза больше, чем при ходьбе босиком (рис.2C) (Chi and Schmitt, 2005; Lieberman et al., 2010). Чтобы избежать этих болезненных и потенциально вредных сил, люди, босые босиком, при беге обычно наносят удары передней или средней частью стопы (рис. 2D). В отличие от ходьбы, бег — это походка массой-пружиной, в которой отсутствуют периоды двойного контакта конечностей, что лишает возможности использовать удар пяткой в энергетике (Cunningham et al., 2010; Perl et al., 2012). Удары передней и средней частью стопы включают приземление с более согнутой лодыжкой, чем удары пяткой, что позволяет контролировать тыльное сгибание голеностопного сустава, что позволяет людям избегать пиков удара тремя способами: за счет уменьшения эффективной массы конечности, которая быстро замедляется, за счет увеличения податливости нижних конечностей и за счет преобразования поступательная энергия, генерируемая при ударе, в энергию вращения в голеностопном суставе (Либерман и др., 2010). Таким образом, люди, по-видимому, способны смягчить высокие ударные силы, связанные с двуногим бегом, посредством кинематических корректировок, а не полагаясь на анатомические адаптации.
Отталкивание и толчок
Будучи двуногими стопоходящими, люди используют стопу для создания значительной положительной силы в голеностопном суставе во второй половине фазы стойки как при ходьбе, так и при беге (Farris and Sawicki, 2012). Для эффективного толчка необходимо укрепить суставы средней части стопы, которые представляют собой суставы между пяточной костью и плюсневой костью (рис.1А) перед высокими внешними моментами. На протяжении десятилетий считалось, что жесткость средней части стопы обеспечивается продольной аркой (см. Глоссарий), уникальной особенностью человека (Рис. 1). Продольная дуга определяется костями среднего отдела стопы, которые удерживаются на месте связками, мышцами и, в первую очередь, подошвенным апоневрозом (см. Глоссарий), широким слоем связочной соединительной ткани. Эта сложная структура прикрепляется проксимально к пяточному бугру и дистально к основаниям проксимальных фаланг, а также к структурам мягких тканей под головками плюсневых костей.Из-за этого дистального прикрепления моменты тыльного сгибания в плюсне-фаланговых суставах (см. Глоссарий) преобразуются в линейную силу, которая напрягает подошвенный апоневроз, приводя в действие «механизм брашпиля» (см. Глоссарий) стопы (рис. 3A, B) (Hicks, 1954). Это натяжение тянет пяточную кость к головкам плюсневых костей, эффективно поднимая продольную дугу и укрепляя среднюю часть стопы во время отталкивания. В своем основополагающем исследовании двуногой ходьбы у шимпанзе Эльфтман и Мантер (1935a) отметили, что, в отличие от людей, средняя часть стопы шимпанзе на короткое время сохраняет контакт с землей после подъема пятки — «разрыв средней плюсны» (см. Глоссарий), впоследствии наблюдаемый у других крупных обезьяны (рис.1B) (D’Août et al., 2002; DeSilva, 2010). Большинство исследователей полагали, что отсутствие разрыва средней предплюсны у людей связано с пассивным усилением продольной дуги через механизм лебедки, и поэтому человекообразные обезьяны не могут придать жесткость средней части стопы во время ходьбы (Bojsen-Møller, 1979).
Рис. 3.Брашпиль-механизм стопы человека. (A) Подошвенный апоневроз прикрепляется проксимально к пяточной кости и дистально к проксимальным фалангам, а бугристость ладьевидной кости указывает высоту продольной дуги.(B) Во время отталкивания тыльное сгибание плюснефаланговых суставов и подошвенный апоневроз обвивают головки плюсневых костей, создавая напряжение, которое создает линейную силу, которая тянет пяточную кость вперед и эффективно поднимает продольную дугу. Это делает ступню жестким рычагом для эффективной передачи усилия от голеностопного сустава. (C) Недавние исследования показывают, что внутренние мышцы стопы (отмечены красным на диаграмме) усиливают механизм брашпиля, активно укрепляя среднюю часть стопы. Эти механизмы позволяют суставам средней части стопы генерировать энергию, которая способствует отталкиванию от голеностопного сустава.
Модель брашпиля функции стопы человека была недавно усовершенствована, чтобы включить в нее вклад внутренних мышц стопы (см. Глоссарий) (рис. 3C). Многие из этих мышц прикрепляются проксимально к подошвенной поверхности пяточной кости и дистально к пальцам стопы и, таким образом, должны быть способны противостоять сжатию продольной дуги при сокращении, подобно сопротивлению, обеспечиваемому подошвенным апоневрозом при тыльном сгибании пальцев стопы (Маккеон и др., 2014). В соответствии с этим ожиданием Келли и его коллеги обнаружили, что несколько собственных мышц стопы подошвенно сгибают суставы средней части стопы при электрической стимуляции во время сидения и изометрически сокращаются после подъема пятки во время ходьбы (Kelly et al., 2014, 2015). Эти данные свидетельствуют о том, что жесткость стопы увеличивается за счет активации внутренних мышц стопы, которые помогают противостоять высоким моментам тыльного сгибания в суставах средней части стопы во время отталкивания.
Это понимание помогает объяснить два недавних удивительных открытия, сделанных в результате экспериментальных исследований передвижения больших обезьян. Во-первых, Bates et al. (2013) измерили распределение подошвенного давления во время двуногой ходьбы у людей, бонобо и орангутанга и обнаружили, что некоторые шаги большой обезьяны перекрываются с некоторыми шагами человека по величине пиков давления, которые, как считается, указывают на податливость средней части стопы (см. Также DeSilva et al., 2015). Во-вторых, используя трехмерную (3-D) кинематику, Holowka et al. (2017b) обнаружили лишь небольшие (но существенные) различия между людьми и двумя шимпанзе в общем объеме тыльного сгибания средней части стопы во время периода поддержки одной конечности при двуногой ходьбе, когда пятка поднимается (рис. 4). Эти данные предполагают, что человекообразные обезьяны могут укреплять суставы средней части стопы во время двуногой ходьбы даже без продольной дуги, что подрывает основное предположение о дихотомии между функцией стопы человека и большой обезьяны.Основываясь на измененной модели брашпиля, наиболее вероятным механизмом этого повышения жесткости является активация мышц стопы, крепления и размеры которых в целом аналогичны у человекообразных обезьян и людей (Oishi et al., 2018). Хотя данные электромиографии in vivo в настоящее время отсутствуют, мы прогнозируем, что собственная мышечная активность стопы тесно коррелирует с жесткостью стопы во время ходьбы у человекообразных обезьян. Если это так, то человекообразные обезьяны могут превращать ступню в умеренно жесткий рычаг во время наземного передвижения, а также поддерживать подвижные суставы средней части стопы для захвата древесной опоры (Holowka et al., 2017а). Тем не менее, человекообразные обезьяны, по-видимому, не в состоянии напрячь ноги так сильно, как люди, во время двуногой ходьбы (Bates et al., 2013; Holowka et al., 2017b), что потенциально ограничивает силу, которую их мышцы могут передавать на землю во время отталкивание, хотя это еще предстоит измерить.
Рис. 4.Движение средней части стопы у человека и шимпанзе при двуногом ходьбе. Полное движение в сагиттальной плоскости для суставов средней части стопы между пяточной костью и плюсневой костью во время фазы стойки у (A) шимпанзе и (B) человека.Углы тыльного сгибания положительные, а углы подошвенного сгибания отрицательные. Вертикальные пунктирные линии обозначают начало и конец поддержки одной конечности. (C) Средний диапазон движений при полной стойке, первая поддержка двух конечностей (DS1), поддержка одной конечности (SS) и вторая поддержка двух конечностей (DS2). Обратите внимание, что шимпанзе использовали немного, но значительно меньше движений во время SS, что согласуется с немного менее жесткой ногой во время отталкивания. Однако люди использовали значительно больше подошвенного сгибания во время DS2 и общего движения в целом, что предполагает более подвижную среднюю часть стопы во время двуногой ходьбы, чем у шимпанзе.Изменено из Holowka et al. (2017b).
Вторая поправка к модели брашпиля вытекает из признания участков значительной положительной и отрицательной работы в ноге. После подъема пятки центр давления перемещается кпереди под стопой, создавая высокие моменты тыльного сгибания в плюснефаланговых суставах, которым должны сопротивляться мышцы-сгибатели пальцев (Rolian et al., 2009) (рис. 5A). Величина этих моментов должна быть соотнесена с длиной зацепа, которая приблизительно равна расстоянию центра давления от этих суставов при отталкивании и, соответственно, плечу нагрузки силы реакции опоры, действующей на них.В исследовании на людях Rolian et al. (2009) обнаружили, что длина пальца не оказывает заметного влияния на расчетную мышечную силу, необходимую для сопротивления моментам в плюснефаланговом суставе во время ходьбы, но оказывает значительное влияние на расчетную мышечную силу во время бега, поскольку бег влечет за собой гораздо более высокие силы реакции опоры на одну конечность. во время отталкивания. У человекообразных обезьян более длинные пальцы ног, чем у людей (Schultz, 1963), и поэтому мы предполагаем, что они должны создавать еще большие силы сгибателей пальцев во время двуногости.Однако шимпанзе и бонобо ходят с меньшими пиковыми углами тыльного сгибания плюснефаланговых суставов в конце стойки, чем люди (Fernández et al., 2016; Griffin et al., 2010). Соответственно, люди обладают дорсально ориентированными суставными поверхностями на головках плюсневых костей («дорсальный купол»; рис. 1А), что отражает способность их плюснефаланговых суставов выдерживать высокие сжимающие нагрузки в позах с сильным тыльным сгибанием, используемых во время двуногого отталкивания (Susman et al. ., 1984). У человекообразных обезьян, напротив, имеются подошвенно ориентированные плюсневые головы, которые, как считается, лучше подходят для суставных поз, используемых при захвате древесных опор.
Рис. 5.Влияние длины пальца на требуемую силу сгибателя при двуногом движении. Во время отталкивания вокруг плюснефаланговых суставов создается внешний момент как продукт силы реакции опоры (GRF) и ее моментного плеча на сустав ( R ). В типичной человеческой стопе (A) этому моменту противодействует сила ( F flexor ) от длинного сгибателя пальцев (FDL), плечо которого ( r ) очень мало.У раннего гоминина с более длинными фалангами (B), но с аналогичным радиусом плюсневой кости ( r ) этот момент увеличен ( R ′), требуя большей силы ( F ′ сгибатель ) от FDL. Изменено из Rolian et al. (2009).
Новые модели ходьбы человека признают, что плюснефаланговые суставы являются потенциально важными участками работы внутри стопы (Zelik et al., 2015), но недавние результаты показывают, что модель функции ступни человека с брашпилем должна быть изменена, чтобы обеспечить положительную работу на стопе суставы среднего отдела стопы (рис.3С). Многосегментные кинематические модели стопы человека демонстрируют значительное подошвенное сгибание средней части стопы, которое совпадает с тыльным сгибанием плюснефалангового сустава во время отталкивания (например, Takahashi et al., 2017). Согласно оценкам обратной динамики, мощность, генерируемая в суставах средней части стопы, более чем компенсирует потерю энергии в плюснефаланговых суставах и совпадает со всплеском силы голеностопного сустава в конце стойки, что способствует чистой положительной работе, производимой стопой во время отталкивания. (Такахаши и др., 2017; Зелик и Хонерт, 2018). Holowka et al. (2017b) обнаружили, что шимпанзе используют значительно меньше подошвенного сгибания средней части стопы, чем люди в конце стойки (рис. 4), что предполагает меньшую способность генерировать энергию во время отталкивания. Кроме того, количество движений средней части стопы, используемое людьми во время отталкивания, даже больше, чем у шимпанзе во время разрыва средней плюсны, что противоречит представлению о том, что люди ходят более жесткими ногами, чем человекообразные обезьяны, и подчеркивает динамическую подвижность человека. стопа во время движения.Эти данные демонстрируют, что грубые наблюдения за движением средней части стопы при ходьбе не точно характеризуют различия во внутренней кинематике стопы у людей и человекообразных обезьян, и выявляют необходимость в дополнительных экспериментальных данных о кинетике суставов средней части стопы. Однако, основываясь на исследованиях, обобщенных выше, мы предполагаем, что люди производят больше внутренней силы ног во время отталкивания при двуногом движении, чем человекообразные обезьяны.
Все модели, рассмотренные до сих пор, предполагали, что первичной структурой, ответственной за жесткость стопы, является продольный свод, что делает эту структуру доминирующим центром сравнительной биомеханики стопы гоминоидов на протяжении десятилетий.Однако Венкадесан и его коллеги недавно предположили, что поперечная дуга также играет жизненно важную роль в укреплении стопы человека (Venkadesan et al., 2017 препринт; Yawar et al., 2017 preprint). Поперечная дуга определяется строением кубовидной и клиновидной костей, оснований плюсневых костей и перекрута стержней плюсневых костей (рис. 6). В своей модели Венкадесан и его коллеги сравнивают эти кости с тонкой пластиной, свободно скрученной на одном конце в поперечной плоскости, как сложенная долларовая банкнота, и геометрически демонстрируют, что эта форма обеспечивает значительное сопротивление изгибу в сагиттальной плоскости.Они утверждают, что у людей эта форма поддерживается задней большеберцовой мышцей проксимально и поперечной плюсневой связкой дистально, и утверждают, что поперечная дуга обеспечивает большую часть пассивной жесткости человеческой стопы. В поддержку этого мнения они показывают, что человеческие ступни примерно в два раза жестче, чем у обезьян, даже с учетом жесткости, обеспечиваемой подошвенным апоневрозом (Bennett et al., 1989). У человекообразных обезьян также наблюдается поперечный изгиб костей предплюсны и основания плюсны (Drapeau and Harmon, 2013), что частично может быть причиной жесткости их стопы при двуногом ходьбе (Bates et al., 2013; Holowka et al., 2017b). Однако из-за подвижных галлюцинаций человекообразных обезьян их поперечные плюсневые связки не связывают плотно первый луч педали с боковыми лучами, как у людей (Raven, 1936), что, вероятно, ограничивает жесткость, обеспечиваемую поперечной дугой.
Рис. 6.Поперечный свод стопы. (A) Поперечный свод стопы человека (вид со стороны) определяется формой кубовидной, клиновидной костей и плюсневых костей, как показано стрелкой.Каркас стопы модифицирован по материалам Эльфтмана и Мантера (1935a). (B) Схематический вид в поперечной плоскости основания плюсневой кости (серый) и головы (белый) у людей и горилл. Пунктирными линиями обозначены приблизительные ориентации голов и оснований. Обратите внимание на большее смещение в ориентации головы и основания в плюсневых костей 3–5 у людей, что создает более четко выраженную поперечную дугу по сравнению с таковой у горилл. Изменено из Мортона (1922).
Упругое накопление и высвобождение энергии
Хотя традиционные сравнения анатомии стопы человека и ноги большой обезьяны выдвинули идею о том, что человеческая стопа эволюционировала как механизм жесткости для двуногой ходьбы (Elftman and Manter, 1935b), Ker et al.(1987) изменили наше понимание биомеханики стопы, показав, что продольный свод стопы функционирует как энергосберегающая пружина во время бега. Бегущее тело можно смоделировать как подпружиненный перевернутый маятник с центром масс тела, падающим в первой половине фазы опоры, растягивая сухожилия в нижних конечностях, которые накапливают потенциальную энергию по мере сгибания суставов. Во второй половине стойки эти сухожилия отталкиваются при разгибании конечности, помогая перемещать центр масс вверх и вперед, а также экономя механическую энергию.Поскольку люди ходят и бегают в стопоходящих позах, сухожилия, пересекающие суставы стопы, не расположены для хранения и высвобождения энергии, как у других животных (Alexander, 1991). Однако сжатие продольного свода стопы под нагрузкой обеспечивает уникальный механизм экономии упругой энергии за счет растягивания эластичных структур в стопе, которые впоследствии отталкиваются во время отталкивания. Ker et al. (1987) впервые протестировали эту пружинную модель, установив трупные ступни человека в устройство, которое циклически нагружало продольную арку в трехточечном изгибе, используя силы, аналогичные тем, которые испытываются при беге с 4-ю.5 м с −1 (рис.7). Путем количественной оценки движения продольного свода во время этих циклов они подсчитали, что энергия, сэкономленная стопой, составляет примерно 17% энергии, необходимой для поддержки веса тела во время фазы опоры бегового шага.
Рис. 7.Накопление и высвобождение упругой энергии в стопе человека. Кривая сила-перемещение, модифицированная по данным Ker et al. (1987) для трупной стопы при моделируемой нагрузке в середине для человека весом 70 кг, бегущего на 4,5 м с -1 .Нагрузка была приложена к основанию стопы с использованием исполнительного механизма для имитации силы реакции опоры (а) и силы трехглавой мышцы на верхней части стопы (b), в результате чего была получена общая сила сжатия на своде стопы (с). Смещение арки измерялось как смещение привода. При приложении силы арка деформируется (стрелки вверх), а затем отскакивает (стрелки вниз), теряя часть энергии (область между кривыми), но возвращая большую часть энергии (заштрихованная синяя область под кривой).
Хотя эта модель имела большое влияние, она не тестировалась in vivo , пока Stearne et al.(2016) измеряли запасы упругой энергии в ногах человека во время ходьбы и бега в минималистичной спортивной обуви и спортивной обуви с индивидуально подобранными стельками, предназначенными для ограничения продольной деформации свода стопы. На основе механической модели, полученной из Ker et al. (1987), они предсказали количество энергии, запасенной в продольной арке, в «нормальных» условиях обуви по сравнению со «стелькой», и сравнили свои прогнозы с измеренными метаболическими затратами на транспортировку. Они обнаружили хорошее соответствие между наборами данных, когда испытуемые набрали 2 балла.7 м с -1 на ровной поверхности, при этом «стелька» увеличивает расход энергии в среднем на 7% и 6% по сравнению с «нормальной» обувью в прогнозах модели и измерениях метаболических затрат соответственно. Эти затраты равняются экономии энергии из-за деформации и отдачи продольной дуги в «нормальном» состоянии обуви. Хотя эта экономия значительно ниже прогнозируемых Ker et al. (1987), в этом исследовании моделировались силы для скоростей бега, которые были на 67% выше, чем те, которые использовались в Stearne et al.(2016), и поэтому ожидается, что это приведет к большей экономии энергии. Наконец, Stearne et al. (2016) не обнаружили разницы в метаболических затратах при ходьбе в «нормальных» и «стельковых» условиях, что указывает на то, что продольный свод стопы не действует как пружина во время ходьбы.
Недавно McDonald et al. (2016) и Вейджер и Чаллис (2016) использовали 3-D моделирование опорно-двигательный аппарат утверждать, что подошвенный апоневрозом является основным местом хранения упругой энергии во время работы. Кроме того, Kelly et al. (2015) обнаружили доказательства того, что внутренние мышцы стопы также активно растягиваются, поглощая энергию в первой половине стойки во время бега, потенциально накапливая энергию, которая может высвобождаться, когда они сокращаются во второй половине стойки.Эластичное накопление энергии потенциально происходит в центральных сухожилиях этих мышц, во внеклеточном матриксе, окружающем их волокна, и в молекулах тайтина их миофиламентов (Roberts, 2016). Независимо от того, какие ткани стопы действуют как пружины, запасание упругой энергии в стопе зависит от наличия продольного свода, и поэтому мы предполагаем, что лапы большой обезьяны не могут экономить значительную энергию во время бега.
Эволюция стопы человека
Исследования, рассмотренные до сих пор, опровергают представление о том, что ступни большой обезьяны приспособлены в первую очередь для лазания, а ступни человека приспособлены только для двуногой ходьбы.Предыдущие исследования использовали эту дихотомию для реконструкции передвижения ископаемых гомининов, которые демонстрируют мозаику черт ног как хождение с формой двуногого движения где-то вдоль континуума от человекоподобного до обезьяноподобного или ходящего с человеческим двуногием, но в основном отказ от древесного поведения. На основе рассмотренных здесь моделей мы вместо этого предлагаем трехэтапный сценарий эволюции стопы человека после расхождения гомининов от последнего общего предка (LCA; см. Глоссарий) шимпанзе и человека (рис.8). (1) Во-первых, в целом большая обезьяноподобная ступня с некоторыми приспособлениями для большей экономичности двуногой ходьбы, но без серьезных компромиссов со способностью хватать педали, что представлено окаменелостями, приписываемыми Ardipithecus ramidus . (2) Во-вторых, нога с приспособлениями для пятки plantigrady и повышенной жесткости средней части стопы во время двуногого шагающего, но сохранил бы некоторую педаль захвата приспособления, как представлено окаменелостями и следами приписываемых австралопитеков афарский .(3) В-третьих, ступня, которая уступила большую часть своей цепкости в пользу приспособлений для способности сохранять упругую энергию и других функций, полезных для ходьбы и бега на большие расстояния, что представлено окаменелостями и следами, приписываемыми ранним представителям этого рода. Человек .
Рис. 8.Трехэтапный сценарий эволюции стопы человека. (A) Стадия 1 представлена окаменелостями Ardipithecus ramidus (Lovejoy et al., 2009) и характеризуется сохранением широко отводящей и противопоставленной большой палец стопы (1) и длинных пальцев стопы (2), но удлинением средней части стопы. (3).(B) Стадия 2 представлена окаменелостями Australopithecus afarensis, (Latimer et al., 1982; Ward et al., 2011) и следами Лаэтоли (пунктирная линия; Leakey and Hay, 1979) и характеризуется пятнистым простиранием пятки. (4) и низкая продольная дуга (5), но сохраняется небольшое отведение галлюцинаций (6) и длинные пальцы ног (7). (C) Стадия 3 представлена окаменелостями Homo habilis и Homo erectus (Day and Napier, 1964; Pontzer et al., 2010), а также следами Илерета (пунктирная линия; Bennett et al., 2009), и характеризуется полностью приведенным большим пальцем стопы (8), продольной аркой, похожей на человека (9), и, судя по отпечаткам пальцев, по-видимому, короткими пальцами (10).
Стадия 1:
Ardipithecus ramidusСамым ранним ископаемым гоминином, у которого были обнаружены обширные останки педалей, является A. ramidus из региона Средний Аваш в Эфиопии (рис. 8A) (Lovejoy et al., 2009 ). Возраст этих окаменелостей составляет 4,4 миллиона лет, что примерно на 2–5 миллионов лет позже LCA (Moorjani et al., 2016). К сожалению, никаких следов, связанных с этим видом, обнаружено не было, и, следовательно, нет прямых доказательств того, что он ходил в стопоходящей позе или использовал удары пяткой, хотя вполне вероятно, что люди и человекообразные обезьяны делают и то, и другое. Тем не менее сохранилось достаточно ископаемого материала, чтобы применить другие модели биомеханики стопы для реконструкции походки A. ramidus . Во-первых, наличие широко расходящегося и противоположного большого пальца стопы, подобного таковому у шимпанзе (Lovejoy et al., 2009), вероятно, указывает на отсутствие продольной дуги, поскольку подвижный большой палец стопы подорвет целостность медиальной стороны дуги, которая является наиболее выраженной областью у людей. Если это так, A. ramidus не смогли бы воспользоваться преимуществом экономии энергии упругости стопы во время бега. Кроме того, как и у человекообразных обезьян, у A. ramidus были длинные изогнутые пальцы, полезные для захвата педали (Richmond, 2007; Susman et al., 1984), но это могло вызвать сильные моменты в плюснефаланговых суставах во время бега (Rolian et al. ., 2009).
Даже при отсутствии продольной дуги, напоминающей человеческую, A. ramidus , вероятно, все еще был способен укрепить свою ступню во время отталкивания с помощью других механизмов, таких как активация собственных мышц стопы (Bates et al., 2013). Кроме того, Lovejoy et al. (2009) утверждали, что A. ramidus мог использовать длинную малоберцовую мышцу для придания жесткости стопе, основываясь на наличии фасетки на кубовиде сесамовидной кости, os peroneum, который обычно встраивается в мышцу. сухожилие у церкопитеков (см. Глоссарий) и гиббонов.Лавджой и др. (2009) утверждают, что эта кость увеличивает плечо рычага сухожилия длинной малоберцовой мышцы и, таким образом, улучшает его способность придавать жесткость средней части стопы. Далее, в значительной степени основываясь на этом механизме, они утверждают, что ступня LCA человека-шимпанзе функционирует аналогично таковой у cercopithecines. Однако этот аргумент подрывается доказательствами того, что гиббоны и церкопитеки имеют очень податливые лапы по сравнению с людьми и шимпанзе (DeSilva, 2010; Greiner and Ball, 2014; Vereecke and Aerts, 2008), а также отсутствием твердых доказательств того, что длинная малоберцовая мышца жесткость средней части стопы у людей.Таким образом, функциональное значение os peroneum неясно. Основываясь на многих сходных с шимпанзе чертах стопы A. ramidus (Pilbeam and Lieberman, 2017), мы предполагаем, что LCA человека-шимпанзе, вероятно, обладал лапой, похожей на обезьянью, как по форме, так и по функциям. .
Тем не менее, некоторые особенности стопы A. ramidus , по-видимому, являются производными приспособлений для двуногой ходьбы, которых нет у человекообразных обезьян. В частности, его кубовидные пропорции указывают на относительно более длинную область средней части стопы, чем у шимпанзе, в пределах диапазона людей (Lovejoy et al., 2009). Если предположить, что A. ramidus использовал позы для стопоходящей ходьбы, удлиненная средняя часть стопы увеличила бы общую длину стопы и, следовательно, расстояние, пройденное центром давления под стопой в фазе стойки. Это могло бы эффективно снизить потерю энергии при столкновении (Adamczyk and Kuo, 2013) и увеличить эффективную длину нижней конечности, чтобы улучшить экономичность ходьбы (Webber and Raichlen, 2016). Кроме того, дорсальное возвышение головки третьей плюсневой кости предполагает, что латеральные (не имеющие большого пальца стопы) плюснево-фаланговые суставы были нагружены в позах с сильным тыльным сгибанием во время ходьбы на двух ногах (Lovejoy et al., 2009; Fernández et al., 2018). Однако голова первой плюсневой кости не имеет дорсального купола, что позволяет предположить, что A. ramidus использовал механику отталкивания, отличную от таковой у людей, возможно, из-за его противопоставления большого пальца стопы.
Стадия 2: австралопитеки
Самое старое свидетельство того, что ступня больше похожа на человеческую, появляется в виде следов, которым 3,66 миллиона лет, с территории Лаэтоли в Танзании, которые чаще всего приписываются австралопитекам (см. ) А.afarensis (рис. 8В) (Leakey, Hay, 1979; Masao et al., 2016). Эти следы характеризуются глубокими отпечатками пятки, что указывает на то, что пятка используется человеком (и шимпанзе) для поддержки веса (Crompton et al., 2012; Hatala et al., 2016a; Raichlen and Gordon, 2017). Они также демонстрируют свидетельства слегка похищенных галлюцинаций по сравнению с обычными босоногими современными людьми (Bennett et al., 2009), но, вероятно, утратившими противопоставимость галлюцинаций человекообразных обезьян (Day and Wickens, 1980).Кроме того, наиболее хорошо сохранившаяся дорожка следа Лаэтоли показывает относительно неглубокую медиальную область средней части стопы, указывающую на продольную арку, хотя в среднем она ниже, чем было измерено в большой выборке современных человеческих следов (Hatala et al., 2016a). Однако, поскольку этот след представляет собой только одного человека и частично пересекается со следами современного человека, эти выводы следует интерпретировать с осторожностью.
Эти выводы из отпечатков ступней подтверждаются несколькими аспектами морфологии костей стопы, сохраненными в 3.Взрослый особь возрастом 2 миллиона лет A. afarensis окаменелостей из формации Хадар в Эфиопии (рис. 8B), а также недавно описанная молодь возрастом 3,3 миллиона лет A. afarensis окаменелость пешком от формации Дикика в Эфиопии (DeSilva et al., 2018). Взрослые окаменелости Hadar демонстрируют большой, похожий на человека пяточный бугорок с умеренно развитым латеральным подошвенным отростком, который некоторые интерпретируют как приспособление для двуногой ходьбы с ударом пятки (Latimer and Lovejoy, 1989; Prang, 2015a), хотя морфологии пяточной кости при рассеивании ударных сил еще предстоит проверить.Кроме того, ориентация суставных поверхностей суставов взрослых и молодых окаменелостей предплюсны и плюсны напоминает ориентацию современных людей и, таким образом, считается показателем продольных и поперечных дуг (DeSilva et al., 2018; Prang, 2015b; Ward et al. др., 2011). Основываясь на этих особенностях, а также на тех, что сохранились на следах Лаэтоли, мы заключаем, что A. afarensis , вероятно, ходил с упором на пятку и использовал некоторую форму поперечных и продольных дуг для придания жесткости ступне.
Морфология галлюкального тарзометатарзального сустава A. afarensis занимает промежуточное положение между суставами человека и человекообразных обезьян по вогнуто-выпуклости суставной поверхности (DeSilva et al., 2018; Gill et al., 2015; Proctor, 2010) , что предполагает наличие большого пальца ноги, который можно было слегка отвести (но см. Latimer and Lovejoy, 1990). Это может указывать на сохранение некоторой способности использовать большой палец стопы для захвата педали во время передвижения по дереву. Это мнение подтверждается морфологией суставной поверхности головки первой плюсневой кости, которая ориентирована подошвенно, как у шимпанзе (Fernández et al., 2016, 2018), что свидетельствует об устойчивости в хватательных позах. Наблюдения за людьми-охотниками-собирателями показывают, что большой палец стопы может быть чрезвычайно полезен для хватания во время лазания, даже при отсутствии противопоставления с другими пальцами (Kraft et al., 2014). Наличие более подвижного большого пальца стопы могло привести к тому, что A. afarensis оттолкнулся от второй и третьей плюсневых головок во время двуногости, а не от первой, как у людей (Fernández et al., 2016), что соответствует относительно неглубоким отпечатки под головкой первой плюсневой кости в следах Лаэтоли (Crompton et al., 2012; Hatala et al., 2016b). Кроме того, A. afarensis сохранил относительно длинные пальцы ног (Susman et al., 1984), что улучшило бы хватание за педали без ущерба для экономики двуногой ходьбы (Rolian et al., 2009). Этот вид обладал похожей на человека морфологией кубовидно-плюсневого сустава, которая, как утверждается, указывает на относительно жесткую среднюю часть стопы, плохо приспособленную для лазания (DeSilva, 2010; Ward et al., 2011). Однако как данные о двуногой кинематике, так и данные о диапазоне движений трупов у людей и шимпанзе не показывают существенных различий между видами в подвижности средней части стопы (Greiner, Ball, 2014; Holowka et al., 2017b), предполагая, что эта особенность не является хорошим индикатором локомоторных возможностей древесных растений.
Эти данные подтверждают мнение о том, что A. afarensis мог ходить на двух ногах с механикой стопы, аналогичной человеческой, сохраняя при этом некоторые приспособления для захвата педали для древесного передвижения. Однако особенности стопы A. afarensis позволяют предположить, что двуногий бег не был важным компонентом локомоторного поведения этого вида. Во-первых, если А. afarensis действительно обладал продольной дугой, она, вероятно, была относительно низкой (DeSilva et al., 2018; Hatala et al., 2016a), и, следовательно, не могла хранить много упругой энергии. Во-вторых, относительно длинные пальцы ноги A. afarensis , вероятно, вызывали высокие моменты в плюснефаланговых суставах во время бега и, следовательно, требовали создания относительно высоких сил мышцами-сгибателями пальцев (Rolian et al., 2009). Хотя бег человека обходится не дороже, чем у других млекопитающих такой же массы (Rubenson et al., 2007), особенности стопы A. afarensis вместе с другими посткраниальными особенностями могли быть приравнены к относительно более высоким метаболическим затратам при беге, которые исключали бы использование бега на длинные дистанции для пропитания (Bramble and Lieberman, 2004) .
Исследования эволюции стопы австралопитеков в основном были сосредоточены на A. afarensis , но несколько других ископаемых гомининов существовали примерно в то же время с морфологически разными ступнями.Нога гоминина возрастом 3,4 миллиона лет из Бёртеле, Эфиопия, демонстрирует противоположный большой палец, как у африканских обезьян, и, следовательно, он мог использовать механику двуногой ходьбы, аналогичную таковой у A. ramidus (Haile-Selassie et al. др., 2012). В Южной Африке стопа Стеркфонтейна возрастом 3,2 миллиона лет, вероятно, Australopithecus africanus , также была описана как имеющая морфологию, указывающую на противоборство большого пальца стопы (Clarke and Tobias, 1995), но эта оценка была оспорена (McHenry и Джонс, 2006).Кроме того, стопа возрастом 2 миллиона лет из Малапы, Южная Африка, отнесенная к Australopithecus sediba , демонстрирует несколько особенностей, которые, как утверждается, влияют на механику двуногой ходьбы, в первую очередь изящную пяточную кость без латерального подошвенного отростка (Zipfel et al., 2011). Утверждалось, что эта кость указывает на то, что A. sediba использовала бы разные позы для удара ногой, чем современные люди, потому что ее пяточка не могла выдерживать высокие силы удара (DeSilva et al., 2013; Prang, 2015a), но, опять же, взаимосвязь между морфологией пяточной кости и сопротивлением переломам еще предстоит проверить. Таким образом, остается неясным, ходил ли A. sediba на двух ногах с механикой стопы, отличной от таковой у A. afarensis .
Стадия 3:
Homo и эволюция ног современного человекаДва геологически одновременных, но географически удаленных участка дали самые ранние ископаемые останки, отнесенные к роду Homo (рис.8С). Стопа возрастом 1,8 миллиона лет из Олдувая в Танзании, обычно относимая к Homo habilis , включает в себя все предплюсневые и плюсневые кости и является убедительным доказательством полностью приведения большого пальца стопы и современной человеческой морфологии сустава средней части стопы (Day and Napier, 1964; Susman and Stern, 1982). Связанные ископаемые плюсневые кости и медиальная клинопись, приписываемая Homo erectus из стоянки Дманиси возрастом 1,8 миллиона лет в Джорджии, также свидетельствуют о полностью приведенном большом пальце стопы, а также о образцах кручения плюсны, указывающих на человеческую поперечную дугу ( Pontzer et al., 2010). Два следа следа возрастом 1,5 миллиона лет из формации Кооби Фора в Кении являются дополнительным доказательством морфологии стопы у Homo (рис. 8C). Размеры отпечатков предполагают гоминины с современными человеческими размерами тела (Behrensmeyer, Laporte, 1981; Dingwall et al., 2013) и, таким образом, отнесены к H. erectus (Bennett et al., 2009). Морфология следов указывает на то, что H. erectus обладал большим приведением большого пальца стопы и продольной дугой, которая была выше, чем на следах Лаэтоли (Bennett et al., 2009; Hatala et al., 2016b), предполагая, что эволюция дуги, похожей на человека, могла произойти за счет способности хватать галлюцинации. Эти следы примерно совпадают с почти полным, но безногим скелетом подростка ( H. erectus ) из соседнего поселения Нариокотоме, который демонстрирует многие анатомические особенности, которые, как утверждается, представляют адаптацию к бегу на выносливость (см. Глоссарий) (Bramble and Lieberman, 2004) . Взятые вместе, эти данные позволяют предположить, что появление высокой продольной дуги, похожей на человека, у H.erectus , возможно, был частично приспособлением для экономии упругой энергии во время бега.
Эволюция более коротких пальцев стопы представляет собой еще один очевидный компромисс между экономией бега и хватанием за педаль (Rolian et al., 2009), но нам еще предстоит обнаружить ископаемые кости пальцев стопы, которые можно твердо отнести к H. erectus или Х. умелый . Неудивительно, что у Homo neanderthalensis пальцы на ногах короткие (Trinkaus, Hilton, 1996), но этот вид появился после первого появления H.erectus более чем на миллион лет. Интересно то, что кости стопы Homo naledi , обнаруженные в пещере Восходящей звезды в Южной Африке, возрастом от 230000 до 330000 лет, сохраняют свидетельства современных человеческих пропорций пальцев ног, но с кривизной фалангового вала, аналогичной кривой горилл (Harcourt- Смит и др., 2015). Утверждается, что эта кривизна указывает на сильную способность хватать за педаль, но наличие полностью приведенной, непротиворечивой большого пальца стопы предполагает, что H. naledi не сохранил значительных древесных приспособлений в своей стопе.В остальном стопа H. naledi очень похожа на современную человеческую, хотя ориентация головки таранной кости и поддерживающей ступни предполагает относительно низкую продольную дугу. Однако связь между этими особенностями и высотой дуги у современного человека невелика и требует дальнейшего тестирования. Таким образом, все производные характеристики стопы современного человека, связанные с энергетической эффективностью во время бега, относятся к роду Homo (за исключением одного непонятного исключения, см. Вставку 1).
Коробка 1.Нечетная стопа Homo floresiensisПочти полный экземпляр стопы (LB1) возрастом от 60 000 до 100 000 лет с острова Флорес приписывается загадочному ископаемому гоминину Homo floresiensis (Sutikna et al., 2016). Этот вид отличается от всех современных гомининов своим удивительно низким ростом и маленьким эндокраниальным размером, а также интересным сочетанием черт, общих с Homo , а также с более ранними гомининами, включая австралопитеков (Brown et al., 2004; Jungers et al., 2009a; Larson et al., 2009). Стопа H. floresiensis не исключение. В целом, лапа LB1 чрезвычайно длинная по сравнению с длиной ноги, отчасти из-за того, что длинные пальцы ног имеют такие же пропорции, как у шимпанзе (Jungers et al., 2009b). Стопа также имеет полностью приведенный большой палец, но его первая плюсневая кость относительно короткая, а кубовидная и ладьевидная кости имеют форму, аналогичную форме африканских обезьян, что позволяет предположить, что продольная дуга была слабо развита или отсутствовала (Jungers et al., 2009b). Эти особенности не препятствуют эффективной двуногой ходьбе, так как длинная ступня должна снизить метаболические затраты, предполагая использование плантиграда с ударом пяткой (Cunningham et al., 2010; Webber and Raichlen, 2016), а для придания жесткости ногам можно было бы использовать альтернативные механизмы. стопа даже при отсутствии продольной дуги (Bates et al., 2013; Venkadesan et al., 2017 препринт). Однако из-за отсутствия продольной дуги, напоминающей человеческую, H. floresiensis не смог бы воспользоваться преимуществом значительной экономии энергии упругости стопы (Stearne et al., 2016), а его длинные пальцы ног потребовали бы высокой мышечной силы во время бега (Rolian et al., 2009). Таким образом, эти особенности несовместимы с адаптацией к бегу на выносливость, что неудивительно, учитывая, что H. floresiensis , вероятно, был эндемиком небольшого лесного острова. Сохранил ли этот вид морфологию стопы от примитивного австралопитекового предка или вторично эволюционировал после заселения Флорес (Jungers et al., 2009b). Изображение скелета стопы, измененное после Jungers et al.(2009b).
Заключительные замечания
Замечательные различия между ногами людей и человекообразных обезьян привели к более чем столетним спорам о том, как и почему человеческая ступня эволюционировала и стала такой, какая она есть. Недавно разработанные и недавно усовершенствованные биомеханические модели помогают нам понять адаптивные функции особенностей, уникальных для человеческой стопы, а также недавние экспериментальные данные исследований передвижения больших обезьян и позволяют нам определить новую основу для понимания эволюции стопы гоминоидов.Мы утверждаем, что вместо простого разделения анатомии стопы человека и большой обезьяны мы должны признать, что у этих видов есть некоторые общие черты, такие как использование стоп стопоходящих позы и способность укреплять суставы средней части стопы во время отталкивания, что было бы полезно для двуногости. в их LCA. Ранние гоминины впоследствии развили такие черты, как зарождающаяся продольная дуга и относительно приведенный большой палец, которые улучшили бы механику отталкивания во время двуногой ходьбы, позволяя сохранить некоторые способности захвата педали.Наконец, с большей приверженностью к наземному движению и опорой на ходьбу и бег на длинные дистанции, гоминины утратили большую часть своих древесных адаптаций в пользу полностью развитой продольной дуги с способностью накапливать упругую энергию.
Недавние доказательства, подтверждающие пружинную модель человеческой ступни при беге, но не при ходьбе, имеют важное значение для понимания адаптивной ценности продольной дуги. Рассмотрение альтернативных механизмов повышения жесткости средней части стопы во время ходьбы (например,г. внутренняя активность мышц стопы, поперечный свод), основным избирательным преимуществом продольного свода может быть обеспечение накопления упругой энергии во время бега на длинные дистанции. Если это так, то основной компромисс при развитии современной человеческой ступни может заключаться не в ходьбе и лазании, а скорее в скалолазании против бега, что соответствует сценарию, предложенному Брамблом и Либерманом (2004) в более общем плане для эволюции посткраниального человеческого тела. анатомия. В будущих исследованиях следует сосредоточить внимание на проверке этого компромисса путем изучения того, как такие характеристики, как высота свода и жесткость у людей, влияют на бег по сравнению с ходьбой и внутренней подвижностью стопы.Кроме того, необходимы дополнительные исследования того, как стопа функционирует в поведении, выходящем за рамки стационарного движения, например, при агрессивных взаимодействиях, ситуациях, требующих быстрого ускорения или маневрировании в сложных условиях. Такое поведение также может иметь большое влияние на физическую форму, и поэтому их изучение имеет решающее значение для определения сил отбора, которые сформировали ногу современного человека.
Благодарности
Мы благодарим двух анонимных рецензентов за полезные отзывы, которые помогли нам улучшить качество статьи.Мы также хотели бы поблагодарить Б. Демеса, П. Фернандеса, В. Юнгерса, С. Ларсона, М. О’Нила, К. Орра и Н. Томпсона за бесценные обсуждения, которые помогли нам осмыслить идеи, изложенные в этой статье.
Сноски
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих или финансовых интересов.
Финансирование
Поддержка этой работы была предоставлена Американской школой доисторических исследований, Фондом Лики и Фондом Веннера-Грена (грант 8836).
- © 2018. Опубликовано ООО «Компания биологов»
Анатомия стопы и лодыжки | Хирургия стопы Луисвилл
Стопа и лодыжка человеческого тела работают вместе, обеспечивая равновесие, стабильность, движение и толчок.
Эта сложная анатомическая структура состоит из:
• 26 костей
• 33 стыка
• Мышцы
• Сухожилия
• Связки
• Кровеносные сосуды, нервы и мягкие ткани
Чтобы понять условия, влияющие на стопу и лодыжку, важно понимать нормальную анатомию стопы и лодыжки.
Лодыжка
Лодыжка состоит из трех костей, прикрепленных мышцами, сухожилиями и связками, которые соединяют ступню с ногой.
В голени находятся две кости, называемые большеберцовой (голени) и малоберцовой костью. Эти кости соединяются (соединяются) с таранной костью или костью голеностопного сустава в тибио-таларном суставе (голеностопном суставе), позволяя стопе двигаться вверх и вниз.
- Большеберцовая кость (большеберцовая кость)
- Фибула
- Талус
- Боковая лодыжка
- Медиальная лодыжка
Костные выступы, которые мы видим и чувствуем на щиколотке:
- Латеральная лодыжка: это наружная кость лодыжки, образованная дистальным концом малоберцовой кости.
- Медиальная лодыжка: это внутренняя лодыжка, образованная дистальным концом большеберцовой кости.
Задняя лапа
Стопу можно разделить на три анатомических части, которые называются задняя, средняя и передняя. Задняя стопа состоит из таранной кости или лодыжки и пяточной кости или пяточной кости. Пяточная кость — самая большая кость стопы, а таранная кость — самая высокая кость стопы. Пяточная кость соединяется с таранной костью в подтаранном суставе, позволяя стопе вращаться в голеностопном суставе.
Задняя лапа соединяет середину стопы с голеностопом в поперечном суставе предплюсны.
Средняя ступня
Средняя ступня содержит пять костей предплюсны: ладьевидную кость, кубовидную кость и 3 клинописные кости. Он соединяет переднюю и заднюю части стопы с помощью мышц и связок. Основная связка — связка подошвенной фасции. Средняя ступня отвечает за формирование сводов стопы и действует как амортизатор при ходьбе или беге.
Средняя ступня соединяется с передней частью стопы в пяти плюсневых суставах плюсны.
- ладьевидная кость
- Кубоид
- Клинопись
Передняя часть стопы
Передняя часть стопы состоит из костей пальцев стопы, называемых фалангами, и плюсневых костей, длинных костей стопы. Фаланги соединяются с плюсневыми костями на подушечке стопы суставами, которые называются фаланговыми плюсневыми суставами. На каждом пальце ноги есть 3 фаланговые кости и 2 сустава, а на большом пальце ноги находятся две фаланговые кости, два сустава и две крошечные круглые сесамовидные кости, которые позволяют пальцу ноги двигаться вверх и вниз.Сесамовидные кости — это кости, которые развиваются внутри сухожилия над костным выступом.
Первая плюсневая кость, соединенная с большим пальцем стопы, является самой короткой и толстой из плюсневых костей и является местом прикрепления нескольких сухожилий. Эта кость важна из-за ее роли в движении и нагрузке.
Анатомия мягких тканей
Наши стопы и лодыжки удерживаются на месте и поддерживаются различными мягкими тканями.
- Хрящ: Блестящий и гладкий хрящ обеспечивает плавное движение в местах, где две кости соприкасаются друг с другом.
- Сухожилия: Сухожилия — это мягкая ткань, которая соединяет мышцы с костями и обеспечивает поддержку. Ахиллово сухожилие, также называемое пяточным канатиком, является самым большим и сильным сухожилием в организме. Расположенный на тыльной стороне голени, он охватывает пяточную или пяточную кость. При воспалении он вызывает очень болезненное состояние, называемое тендинитом ахиллова сухожилия, и может сделать ходьбу практически невозможной из-за боли.
- Связки: Связки представляют собой прочную веревку, подобную ткани, которая соединяет кости с другими костями и помогает удерживать сухожилия на месте, обеспечивая стабильность суставов.Подошвенная фасция — это самая длинная связка стопы, берущая свое начало от пяточной кости, пяточной кости и продолжающейся по нижней поверхности стопы до переднего отдела стопы. Он отвечает за свод стопы и обеспечивает амортизацию. Подошвенный фасциит, частая причина боли в пятке у взрослых, может возникать, когда в подошвенной фасции возникают повторяющиеся микроразрывы в результате чрезмерного использования. Растяжение связок голеностопного сустава, наиболее часто регистрируемое повреждение области стопы и голеностопного сустава, связано с растяжением связок и обычно возникает в таранно-малоберцовой связке и пяточно-малоберцовой связке.
- Мышцы: Мышцы представляют собой фиброзную ткань, способную сокращаться, вызывая движение тела. В стопе 20 мышц, которые классифицируются как внутренние и внешние. Собственные мышцы — это те, которые расположены в стопе и отвечают за движение пальцев ног. Внешние мышцы расположены снаружи стопы в голени. Икроножная или икроножная мышца является самой большой из них и способствует движению стопы. Растяжения мышц обычно возникают из-за чрезмерного использования мышцы, в которой она растягивается без должного разогрева.
- Бурсы: Бурсы — это небольшие мешочки, заполненные жидкостью, которые уменьшают трение между сухожилиями и костью или кожей. Бурсы содержат особые клетки, называемые синовиальными клетками, которые выделяют смазочную жидкость. Когда эта жидкость заражается, может развиться обычное болезненное состояние, известное как бурсит.
Биомеханика стопы и голеностопного сустава
Биомеханика — это термин, описывающий движение тела. Сам по себе голеностопный сустав допускает два движения:
- Подошвенное сгибание: стопа направлена вниз.Это движение обычно сопровождается переворачиванием стопы.
- Тыльное сгибание: подъем стопы вверх. Это движение обычно сопровождается выворотом стопы.
Стопа (без пальцев) также допускает два движения:
- Инверсия: поворот ступни внутрь.
- Eversion: поворот подошвы стопы наружу
Пальцы ног допускают четыре различных движения:
- Подошвенное сгибание: сгибание пальцев стопы к подошве стопы
- Тыльное сгибание: сгибание пальцев стопы к вершине стопы
- Похищение: Разведение пальцев ног.Это движение обычно сопровождает подошвенное тыльное сгибание.
- Приведение: Соединение пальцев ног вместе. Это движение обычно сопровождает подошвенное сгибание.
Ссылки по теме
иллюстраций костей стопы и анатомии стопы
Медицинские иллюстрации для анатомического образования
Мы можем предоставить индивидуальные или стандартные медицинские иллюстрации, демонстрирующие анатомию стопы человека.На протяжении многих лет нам было поручено завершить медицинские иллюстрации для публикации книг по анатомии, плакатов и диаграмм по анатомии, а также на веб-сайтах по санитарному просвещению. В галерее представлены некоторые примеры выполненных нами работ. Однако мы можем получить заказ на создание индивидуальных иллюстраций для любого проекта.
Анатомия стопы
Стопа человека представляет собой специально разработанную сложную структуру, состоящую из 26 костей и 33 суставов, работающих вместе с 19 мышцами и 107 связками, чтобы выполнять высокоточные движения при движении.Во время движения стопа передает вес тела, а также силы, во много раз превышающие вес человеческого тела, для выполнения таких действий, как ходьба, бег и прыжки, и даже способна приспосабливаться, чтобы справляться с различными ландшафтами.
Стопа способна так хорошо двигаться благодаря расположению ее костей, связок и фасций, которые образуют пружинную арочную структуру с толстыми подушечками под пятками и пальцами ног для амортизации и поддержки. Эти структуры, называемые сводами стопы, образованы костями предплюсны и плюсны и укреплены связками и сухожилиями.
Арки стопы
Три арки называются медиальной продольной аркой, расположенной вдоль внутренней стороны стопы, и боковой продольной аркой, которая представляет собой меньшую арку вдоль внешней стороны стопы, и одну переднюю поперечную арку, проходящую через переднюю часть стопы, поскольку о его местонахождении нельзя увидеть в действии. Вместе их форма и структура позволяют им действовать так же, как пружина, выдерживая вес тела и поглощая удары, возникающие во время передвижения.
Анатомические сведения о стопе человека
Саму ступню можно разделить на три отдельные части: заднюю, среднюю и переднюю. Задняя лапа состоит из двух из семи костей предплюсны, таранной кости и пяточной кости. Средняя часть стопы содержит остальные кости предплюсны, а передняя часть стопы — плюсневые кости и фаланги.
Из всех костей стопы таранная, кубовидная и средняя клинопись являются структурно наиболее важными, так как они являются так называемыми тремя замковыми камнями.Самый важный компонент любой арочной конструкции — Keystone. Это кость или «замковый камень», который образует вершину арки и скрепляет всю структуру.
Кости производят клетки крови, называемые гематопиэзом, плюс они накапливают минералы в костном матриксе. Такие, как кальций, участвуют в метаболизме кальция, а костный мозг может хранить железо в ферротине и участвует в метаболизме железа. Костная структура состоит из смеси хондроитинсульфата и гидроксиапатита, составляющих 70% кости.
футов: анатомия и физиология | Encyclopedia.com
Человеческая стопа — это одновременно самая функциональная, самая сложная и самая опасная часть анатомии. Сложную скелетную структуру стопы сравнивают с сочетанием механизмов и долговечностью прекрасных швейцарских часов — инструмента, способного перемещать тело в любом направлении с высокой скоростью, выдерживая при этом силы, во много раз превышающие вес тела человека. Как структура, которая поддерживает тело во всех аспектах вертикального движения, стопа подвергается постоянным физическим нагрузкам, некоторые из которых усугубляются такими условиями, как обувь и погода.Как и многие другие компоненты анатомии человека, которые функционируют постоянно, стопа замечается только тогда, когда она перестает нормально функционировать.
Стопа превратилась в течение сотен тысяч лет физиологического развития человека в механизм, который является одновременно сложной машиной и основой тела. Включая кости лодыжки, ступня состоит из 26 отдельных костей, многие из которых относительно небольшие и имеют изящную форму. Более 100 мышц, сухожилий и связок в сочетании со скелетными костями образуют 33 отдельных сустава в этой гибкой и динамичной структуре.Сложная серия кровеносных сосудов и нервных путей проходит в отдельных сетях, окруженных тканями стопы, чтобы поддерживать сердечно-сосудистые и неврологические потребности в движении. Сложность скелетной структуры стопы подчеркивается тем фактом, что кости двух человеческих стоп составляют почти 25% всех костей человеческого тела.
Неравномерность или дисбаланс в структуре стопы часто приводит к поражению других анатомических структур.Например, если человек имеет тенденцию с силой ударяться о землю передней частью подошвы стопы, а не пяткой, большая степень силы имеет тенденцию распространяться на коленный сустав или бедро.
Костная структура стопы разделена на три части: переднюю часть стопы, среднюю часть стопы и заднюю стопу. Передняя часть стопы состоит из костей пяти пальцев стопы, которые вместе известны как фаланги. Фаланги соединены с другими костями стопы более длинной соединительной костью, называемой плюсневой костью, в суставах, образованных на подушечке стопы с каждым пальцем.Передняя часть стопы способна выдерживать половину веса человека.
Средняя часть стопы — это часть стопы, которая предназначена для поглощения ударов, создаваемых движением человека. Середина стопы состоит из пяти костей предплюсны и поддерживается подошвенной фасцией, связкой, которая важна для функции свода стопы. Подошвенная фасция проходит по всей длине стопы и прикрепляется к пяточной кости (пяточная кость, самая большая кость стопы) к передней части стопы.Задний отдел стопы, включая структуру голеностопного сустава, соединен с костями голени таранной костью, голеностопной костью. Соединение, созданное на пятке и голеностопном суставе, является подтаранным суставом, который позволяет голеностопному суставу полностью вращаться по часовой стрелке и против часовой стрелки.
Двадцать мышц, которые вызывают движение в стопе, имеют такую же тонкую и сложную структуру, как и скелетная кость стопы-компаньона. Эти мышцы, как и ахиллово сухожилие, ответственны за создание стопой всех типов движений.Например, передняя большеберцовая мышца позволяет ступне двигаться вверх, что необходимо для отрыва передней части стопы от земли. Задняя большеберцовая мышца — это мышца, поддерживающая арку. Малоберцовая большеберцовая мышца контролирует движение на внешней стороне лодыжки, например поворот стопы на ее внешний край. Разгибатели используются, чтобы помочь лодыжке поднять пальцы ног, когда тело готовится к шагу вперед. Мышцы-сгибатели стабилизируют пальцы ног на земле, особенно когда тело неподвижно и вертикально.Ахиллово сухожилие, которое соединяет пятку с икроножной и камбаловидной мышцами, или икроножными мышцами, является самым большим и сильным сухожилием в организме. Ахиллес необходим для обеспечения стабильности всей анатомической структуры голени при беге или прыжке.
Связки стопы образуют суставы, способные значительно сгибать пищу и выдерживать большой вес. Подошвенная фасция — самая длинная из связок стопы, действующая как амортизирующее устройство для всей конструкции во время движения.У каждого пальца ноги есть маленькие суставы, образованные связками, чтобы обеспечить гибкость каждого из этих придатков, независимо от остальной структуры.
Каждое движение стопы представляет собой серию синхронизированных мышечно-скелетных движений, совершаемых в координации с голеностопом и голенью. Каждое движение тела, связанное с его движением вперед, назад или вверх, осуществляется этими компонентами комплексно. В спорте, когда стопа находится в неподвижном положении, она остается функциональной.Примером этого состояния является гребля, где основной упор спортсмена делается на выполнение гребка весла в воде, а ступня постоянно сгибается и способствует общему равновесию и устойчивости тела в плавсредстве. При занятиях такими видами спорта, как стрельба и парусный спорт, неподвижные ступни спортсмена способствуют эффективному выполнению спортсменом других физических задач.
см. Также Разрыв ахиллова сухожилия; Анатомия и физиология голеностопного сустава; Анатомия голени.
